
Projekt Pechgraben
News & Aktuelles
_____________________________________________________________________________________________
Herausgeber / Editors: Dipl.-Biol. Andreas Hecker & Dr. Alexander M. Heyng
_____________________________________________________________________________________________
- Aktuelle Publikation -



click to enlarge & read figure captions
Publication:
Bicknell RDC, Hecker A, and Heyng AM. (2021) New horseshoe crab fossil from Germany de-monstrates post-Triassic extinction of Austrolimulidae. Geological Magazine
https://doi.org/10.1017/S0016756820001478
Link zur Pressemitteilung der SNSB vom
15. Februar 2021:
https://snsb.de/neuer-alter-pfeilschwanzkrebs-aus-franken/
A team of palaeontologists discovered a 197 million year old Jurassic horseshoe crab in Franconia, Bavaria. In considering this fossil, they came to a shocking conclusion: the horseshoe crab belonged to a family that was thought to be extinct at the end of Triassic.
The material was gathered from a locality called Pechgraben, near Neudrossenfeld (Franconia, Bavaria, Germany). These rocks are considered to come from the Bayreuth-Formation (Hettangium, ~197 million years old). In particular, rare clay lenses at this locality have preserved numerous and valuable plant fossils, but just a few animal fossils. However, in 1999, the Paläontologisches Museum Nierstein conducted an excavation and uncovered a 5 cm long horseshoe crab fossil; a fossil that could not easily be placed into other known species.
This interesting fossil became the focus of Russell Bicknell (University of New England, Armidale, Australia), Andreas Hecker (Jura-Museum Eichstätt) and Alexander M. Heyng (amh-Geo). Examination of the fossil determined that it belonged within its own genus, and was from the family Austrolimulidae. This outcome was a shock, as Austrolimulidae is traditionally considered to have gone extinct over the Triassic-Jurassic-boundary 201 million years ago. The research team named the fossil Franconiolimulus pochankei in honour of the region Franconia and Hartmut Pochanke, the sand pit worker who discovered the clay deposits and, by extension, allowed this fossil to be identified.
Horseshoe crabs are very peculiar animals. Contrary to their common name, they are no crustaceans. In fact they are more closely related to spiders and scorpions. Even more impressive: they are alive today! The most iconic is the American horseshoe crab Limulus polyphemus which is observed along the west coast of America, between Mexico and Maine. Their overall shape - a body consisting of two sections and a long and slender tail (telson) - has not changed dramatically over the last several hundred million years. All modern horseshoe crabs belong to Limulidae - a family that appeared in the mid Triassic, ~ 240 million years ago.
To understand the life mode of Franconiolimulus pochankei, the paleoenvironment in which the new horseshoe crab lived was reconstructed through discussions with colleagues at the Urweltmuseum Oberfranken. It seems that ~197 million years ago, Franconiolimulus lived in a tropical river channel surrounded by large fern trees, giant horsetails, and gymnosperms. In considering this paleoenvironment, it was also identified that animal fossils are much rarer than previously thought. However, this is not overly surprising as it is similar to modern tropical streams where animal carcasses rapidly decay.
- Published online 22-02-2021 by Bicknell, Hecker & Heyng
-
_____________________________________________________________________________________________
- Publikationen zum Thema -
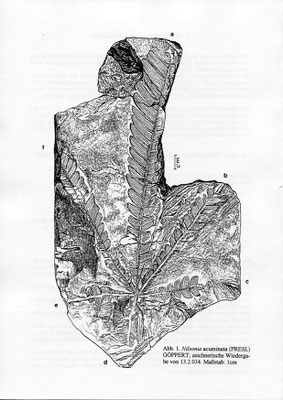
Ein Blattschopf von Nilsonia acuminata (PRESL) GOEPPERT
aus dem Unteren Lias Oberfrankens
von St. Schmeissner & S. Hauptmann
Zeitschrift Documenta naturae Bd. 117 / 1998
Zusammenfassung
Aus dem Unteren Lias Oberfrankens wird ein Blattschopf von Nilsonia acuminata beschrieben. Der Fund beweist, daß die Blätter zu mehreren in Büscheln, vermutlich an Kurztrieben angeordnet waren. Rekonstruktionsversuche mit cycadeen-ähnlichem Habitus sind somit widerlegt.
Einleitung
Von den weltweit ca. 120 Nilsonia-Arten, die aus dem Zeitraum der Oberen Trias bis zur Unteren Kreide stammen, sind im Unteren Lias Frankens
folgende Arten nachgewiesen:
N. acuminata
N. minima
N. polymorpha
N. „typ “ orientalis (es kann sich dabei auch um eine Pflanze aus der Formgattung Taeniopteris oder um Nilssoniopteris handeln (frdl. mündl. Mitteilung von
Dr. VAN KONIJNENBURG - VAN CITTERT).
Bei den meisten Arten sind die Blattspreiten in Fiedern unterteilt; diese sind paralleladrig, sehr selten treten einfache Dichotomien auf. Kennzeichen aller Nilsonien (der Gattungsname
Nilsonia ist aus Prioritätsgründen richtig und nicht die weit verbreitete Schreibweise Nilssonia (vgl. KELBER & HANSCH: 128) ist die Anheftung der Fiedern auf der Oberseite
der Rhachis, die dadurch von oben betrachtet verdeckt wird.
Als weitere den Nilsonien zugerechnete Organe findet man Samen, männliche Blüten (Androstrobus\ weibliche Blüten (Beania) bzw. Samenstände, Pollen und unter dem Namen
Deltolepis schuppenförmige Blättchen (Bracteen). Aus den fränkischen Fundgebieten sind neben den Blättern bislang nur evtl, zugehörige Samen beschrieben worden..
Historisches
Obwohl Nilsonien im Mesozoikum weit verbreitet waren und z.B. im Unteren Lias Frankens zu den häufigen Fossilien gehören, ist über die Wuchsform dieser Pflanzenfamilie lange Zeit nichts Gesichertes bekannt geworden. Hinweise auf eine schopf-förmige Anordnung der Blätter geben Funde mit einem entsprechenden Arrangement. Da die eigentliche Blattbasis überall fehlt, ist die Aussagekraft solcher Stücke stets eingeschränkt. Folgende Beispiele seien aufgefuhrt:
N. acuminata: MÄGDEFRAU (1968: 306, Abb. 253a)
N. polymorpha: NATHORST (1909: Taf. 4 und Taf. 6, Abb. 1 u. 2)
Eine ausführliche Liste findet sich bei KIMURA & SEKIDO (1975: 115 f.).
Rekonstruktionsversuche zeigen Nilsonien meist als sehr cycadeenähnlich: An der Spitze eines unverzweigten, aber stets imaginären Stammes sitzt der Blattschopf (vgl. PROBST, E. (1986: 147) und GOTHAN (1926) in MÄGDEFRAU (1968: 320, Abb. 265)).
Gegen solche Vorstellungen wandte sich bereits NATHORST: „Ich halte es für wenig wahrscheinlich, dass Nilssonia knollen-förmige oder zylindrische Stämme gehabt hat, vielmehr könnte man vermuten, dass die Stämme wiederholt gegabelt waren (aus: NATHORST 1909: 30).
Eine entsprechende Rekonstruktion der mitteljurassischen N. tenuinervis gibt HARRIS (1961: 321): An mindestens 2cm dicken, sich vielleicht wiederholt gabelnden Stämmen, sitzen an den Enden spiralig verteilt die Blätter. HARRIS betont jedoch, daß der Stamm frei erfunden ist.
Fundort und Geologie
1990 entdeckte einer der Autoren (S. Hauptmann) den im folgenden beschriebenen Schopf von Nilsonia acuminata. Das Stück befindet sich unter der Registriemumnier 13.2.034 in der Sammlung Hauptmann / Hof.
Die Fundstelle liegt im östlichen Landkreis Kulmbach, im Bereich des obermainischen Bruchschollenlandes. Der hier anstehende fluviatile Sandstein des Unteren Lias wird in zahlreichen Sandgruben abgebaut. In den Sandstein eingelagerte Tonlinsen liefern immer wieder gut erhaltene Pflanzenreste, meist als Abdruck, gelegentlich noch mit kohliger Substanz.
Der Blattschopf stammt aus einer Tonlinse der Sandgrube Küfner, südlich der Ortschaft Pechgraben gelegen (TK 1 : 25000, Blatt 5935 Marktschorgast, R 44 67225, H 55 40780).
Dieselbe Tonlinse lieferte von 1988 bis 1994 weitere interessante Pflanzenreste, über die bereits Publikationen vorliegen: KIRCHNER (1992), KIRCHNER & VAN KONUNENBURG - VAN CITTERT (1994) und SCHMEISSNER & HAUPTMANN (1993).
Beschreibung des Materials und Rekonstruktion
Nilsonia acuminata (PRESL in STERNBERG) GÖPPERT 1844
Abb. 1 und 2; Tafel 1, Fig. 1 und Tafel 2, Fig. 2
1838 Zamites acuminatus - PRESL in STERNBERG; S. 199, Taf. 43, Fig. 2
1838 Zamites heterophyllus - PRESL in STERNBERG; S.199, Taf. 43, Fig. 4 u. 5
1844 Nilssonia acuminata - GÖPPERT; S. 141
1867 Nilssonia acuminata - SCHENK; S. 131; Taf. 32, Fig. 1 bis 7
1914 Nilssonia acuminata - GOTHAN; S.35 ff.; Taf. 26, Fig. 1; Taf. 28, Fig. 2 u. 3; Taf. 30,
Fig. 2 bis 4; Taf. 31/32, Fig. 2 u. 3; Taf. 33, Fig. 4; Taf. 34,
Fig. 2 u. 4; Taf. 38, Fig. 2 bis 5; Taf. 39, Fig. 1 u. 2
1966 Nilssonia acuminata - URLICHS; S. 27
1968 Nilssonia acuminata - WEBER; S. 55; Taf. 11, Fig. 111; Taf. 12, Fig. 118 bis 120
Die aus fünf Teilen zusammengesetzte Liegendplatte (die Hangendplatte zersplitterte bei der Bergung vollständig) zeigt das als Abdruck in feinkörnigem grauen Ton erhaltene Fossil von der
Blattoberseite, denn die einzelnen Segmente (Fiedern) sind entlang der Mittellinie an der Oberseite der Blattachse befestigt. Diese ist daher verdeckt.
Bei allen sechs Blättern ist die jeweilige Basis erhalten; deutlich erkennbar ist ihr gemeinsamer Ausstrahlungspunkt (Taf. 1, Fig. 1 u. Taf. 2, Fig. 2; Abb. 1).
Von dem ursprünglich im Zentrum vermutlich vorhandenem Kurztrieb ist jedoch keine Spur mehr zu sehen.
Blatt a ist mit 18cm Länge am vollständigsten erhalten; seine Breite beträgt knapp 3 cm. Blatt d ist mit 3,5cm am kürzesten.
Die schräg ansitzenden Blattsegmente sind stets bis zur Rhachis getrennt. Ihre Länge nimmt von der Blattbasis an gleichmäßig zu; vollständig erhaltene Einzelblätter zeigen zur Spitze hin wieder eine Längenreduktion der Fiedem. Die Breite der Segmente vergrößert sich dagegen nur unwesentlich.
Die Form der Fiedern ist recht variabel; sie reicht von rundlich über dreieckig bis hin zu annähernd viereckig. Besonders bei den schmalen Segmenten sind die Spitzen mehr oder weniger deutlich sichelförmig zur Blattspitze hin gekrümmt. Die Aderndichte der parallelen, ungeteilten Adern beträgt etwa 2 / mm.
click to enlarge & read figure captions



Bei der Rekonstruktion (Abb. 2) wurden die Blätter auf Größen zwischen 12 und 21cm ergänzt. Dabei orientierten wir uns an vollständig erhaltenen Einzelblättem. Diese hatten zwar gelegentlich Ausmaße bis ca. 50cm, jedoch erreichte keines der Einzelblätter unseres Schopfes eine solche Dimension. Das größte Blatt dürfte eine Länge zwischen den angegebenen 21cm und maximal 30cm gehabt haben. Die unterschiedliche Länge der einzelnen Blätter ergibt sich aus der verschiedenen Breite und Breitenzunahme der jeweiligen Segmente.
Die Anzahl der Blätter erhöhten wir auf sieben, da sich aufgrund des größeren Abstandes zwischen Blatt c und d dort ursprünglich noch ein weiteres befunden haben könnte.
Diskussion
Den bereits eingangs angeführten Beispielen einer schopftormigen Anordnung von Nilsonia-Blättern kann an dieser Stelle ein weiters zugefügt werden.
Frau Dr. J. H. A. VAN KONIJNENBURG - VAN CITTERT stellte uns das Foto einer Nilsonia compta aus dem Mittleren Jura Yorkshires zur Verfügung (Taf. 2, Fig. 3). Die drei erhaltenen Blätter, von denen zwei sich in Richtung ihrer Basis zu überdecken, laufen auf ein gemeinsames Zentrum zu. Da die Blattbasen jedoch nicht erhalten sind, ist es unmöglich zu beweisen, ob die Blätter so eng zusammen standen wie bei dem Fund aus Pechgraben. Vielleicht bildeten sie einen breiten Schopf, vergleichbar dem der rezenten Cycadeen.
Die vermutlich treffendste Anschauung für die Wuchsform von N. acuminata liefert Nilssoniocladus nipponense (YOKOYAMA) KIMURA & SEKIDO
1975: An einem 30cm lang erhaltenen und 6mm breiten Langtrieb sind im Abstand von 5 bis 6cm Kurz-triebe spiralig inseriert. Diese sind 1,5cm lang und 4 bis 5mm breit; an der Spitze sitzt jeweils
ein Schopf bestehend aus 3 bis 7 Blättern der Art Nilsonia nipponense. Der bei KIMURA & SEKIDO (1975, Taf. 2, Fig. 1) wiedergegebene Blattschopf zeigt die gleiche Situation wie der
von 13.2.034 (s.Taf. 1, Fig. 1 u. Taf. 2, Fig. 2): Der in der Horizontalen vollständig ausgebreitete Schopf ist von der Oberseite sichtbar. Falls der Kurztrieb noch vorhanden ist, steckt er im
Zentrum senkrecht im Gestein.
Die Wuchsform betreffend, meinten die Autoren, daß es sich bei Nilssoniocladus nipponenese um einen Baum oder möglicherweise um eine Kletterpflanze handelt (ebd.: 112). In Bezug auf
andere Nilsonien schreiben die Autoren: „...but we know of a considerable number of rock slabs which show a group of leaves of which the base was unfortunately lost in collecting. These radiate
from a point, suggesting a cluster as in Nilssoniocladus and it is thus likely that other species have very similar stems“ (aus: KIMURA & SEKIDO 1975: 115).
Aufgrund des in dieser Arbeit beschriebenen Fundes trifit diese Vermutung auf N. acuminata ganz besonders zu, und wir dürfen einen recht ähnlichen Habitus für sehr wahrscheinlich erachten.
Daß KIMURA & SEKIDO die Wuchsform von Nilssoniocladus richtig einschätzten, zeigen neue Funde aus der Kreide von Alaska und Rußland. SPICER & HERMAN (1996) belegen für Nilssoniocladus alaskensis eine große Ähnlichkeit in der Organisation der Lang- und Kurztriebe mit Ginkgo biloba. Bei Nilssoniocladus chukotensis berichten die Autoren von einer schopfiormigen Anordnung der Blätter an Kurztrieben.
Dank
Unser Dank gilt allere die uns bei der Erstellung dieser Arbeit unterstützt haben. Namentlich genannt seien Dr. J. H. A. Van KONUNENBURG - VAN CITTERT für ihre wertvollen Ratschläge , K.-P. KELBER für die Anfertigung von Fotografien, sowie A. LANGOLD und R. GAGEL für ihre Hilfen am Computer.
Schriftenverzeichnis
BARNARD, P.D.W. & MILLER, J. (1976): Flora of the Shemshak Formation (Elburz, Iran), part 3: MiddleJurassic (Dogger) Plants from Katumbargah, Vasek Gah
and Imam Manak. - Palaeontographica B, 155, 1-4: 31-117; Stuttgart.
BENDA, L. (1964): Die Jura - Flora aus der Saighan - Serie Nord - Afghanistans. - Beih. Geol. Jb. 70: 99 -152; Hannover.
FLORIN, R. (1920): Über den Bau der Blätter von Nilssonia polymorpha SCHENK. - Ark. Bot. 16, 7: 1 - 10; Uppsala.
GOTHAN, W. (1914): Die unterliassische (rhätische) Flora der Umgegend von Nürnberg. - Abh. Naturhist.Ges. Nürnberg. - 19: 89 - 186; Nürnberg.
GOTHAN, W. & WEYLAND, H. (1964): Lehrbuch der Paläobotanik. - 2. Aufl.: 593 S., Akademie - Verlag; Berlin.
HALLE, G. (1913): The mesozoic flora of Grahamland. - In: Wissenschaftliche Ergebnisse der schwedischen Südpolar - Expedition 1901 - 1903; 3, 14: 119 S.; Stockholm.
HARRIS, T. M. (1932): The fossil flora of Scoresby Sound, East Greenland. Part 2: Description of seed plants incertae sedis together with a discussion of certain cycadophyte cuticles. - Medd. om
Grönland, 85 (3): 1-114; Kobenhavn.
HARRIS, T. M. (1942): Notes on the Jurassic Flora of Yorkshire. 2. Deltolepis crepidota gen. et sp. nov.; 3. On Nilssonia compta and its reference to Beania
gracilis. - Ann. Mag. Nat. Hist. (11), 9: 573 - 587; London.
HARRIS, T. M. (1961): The fossil cycads. - Palaeontology, 4 (3): 313 - 323; London.
HARRIS, T. M. (1964): The Yorkshire Jurassic Flora, Vol. 2. Caytoniales, Cycadales & Pteridosperms. - 191 S., British Museum (Natural History); London.
JOHANSSON, N. (1922): Die rätische Flora der Kohlengruben von Stabbarp und Skromberga in Schonen. - Kungl. Svenska Vetensk. Akad. Handl. 63, 5:1 - 78; Stockholm.
KELBER, K. - P. & HANSCH, W. (1995): Keuperpflanzen. Die Enträtselung einer über 200 Millionen Jahre alten Flora. - 157 S., museo 11; Heilbronn.
KIMURA, T. & SEKIDO, S. (1975): Nilssoniocladus n. gen. (Nilssoniaceae n. fam.), newly found from the Early Lower Cretaceous of Japan. - Palaeontographica B, 153: 111-118;
Stuttgart.
KIRCHNER, M. (1992): Untersuchungen an einigen Gymnospermen der fränkischen Rhät-Lias-Grenzschichten. - Palaeontographica B, 224, 1-3: 17-61; Stuttgart.
KIRCHNER, M. & VAN KONIJNENBURG - VAN CITTERT, J. H. A. (1994): Schmeissneria microstachys (PRESL 1833) KIRCHNER & VAN KONIJNENBURG -VAN CITTERT comb. nov. and Karkenia
hauptmanni KIRCHNER & VAN KONIJNENBURG - VAN CITTERT sp. nov., plants with ginkgolean affinities from the Liassic of Germany. - Rev. Palaeobot. Palynol., 83, 1 -3: 199 - 215;
Amsterdam.
MÄGDEFRAU, K. (1968): Paläobiologie der Pflanzen, - 4. Auf!.: 549 S., G. Fischer Verlag; Stuttgart.
NATHORST, A G. (1897): Zur mesozoischen Flora Spitzbergens. - Kongl. Svenska Vetensk. Akad. Handl. 30, 1:1 - 77; Stockholm.
NATHORST, A. G. (1902): Beiträge zur Kenntnis einiger mesozoischer Cycadophyten. - Kungl. Svenska Akad. Handl., 36, 4: 25 S.; Stockholm.
NATHORST, A. G. (1909): Über die Gattung Nilssonia BRONG. mit besonderer Berücksichtigung schwedischer Arten. - Kungl. Svenska Akad. Handl. 43, 12: 1 - 40; Stockholm.
PROBST, E. (1986): Deutschland in der Urzeit. - 479 S. Bertelsmann; Stuttgart.
REYMANOWNA, M. (1963): The Jurassic Flora from Grojec near Cracow in Poland. Part 1. - Acta Palaeobot., 4, 2: 9-48; Krakau.
SCHENK, A. (1867): Die fossile Flora der Grenzschichten des Keupers und Lias Frankens. - 232 S. & Atlas; Würzburg.
SCHMEISSNER, S. & HAUPTMANN, S. (1993): Fossile Pflanzen aus den Rhät - Lias - Übergangsschichten des Kulmbach - Bayreuther Raumes. „Glossophyllum“ - ein 200 Millionen Jahre alter
Baum. - Naturwiss. Ges. Bayreuth, Ber. XXII: 51-66; Bayreuth.
SPICER, R. & HERMAN, A. (1996): Nilssoniocladus in the Cretaceous Arctic: new species and biological insights.- Rev Palaeobot. Palynol., 92: 229 - 243; Amsterdam.
STEWART, W. N. & ROTHWELL, G. W. (1993): Palaeobotany and the evolution of plants. - 2. Aufl., 521 S., Cambridge University Press; Cambridge.
URLICHS, M. (1966): Zur Fossilfiihrung und Genese des Feuerlettens, der Rät - Lias - Grenzschichten und des unteren Lias bei Nürnberg. - Erlanger geol. Abh. 64: 1 - 42; Erlangen.
WEBER, R. (1968): Die fossile Flora der Rhät - Lias - Übergangsschichten von Bayreuth (Oberfranken) unter besonderer Berücksichtigung der Coenologie. - Erlanger geol. Abh. 72: 1 -73;
Erlangen.
- Published online 19-12-2023 by A.M. Heyng -
Neue Ergebnisse zu Ctenozamites wolfiana (Pteridospermae)
und Pseudoctenis prossii nov.spec. (Cycadophyta)
aus dem Unteren Lias (Jura, Bayern)
von
J.H.A. Van Konijnenburg - Van Cittert, St. Schmeissner, S. & T. Hauptmann
Zeitschrift Documenta naturae Bd. 117 / 1998
Zusammenfassung
Einige neue Fragmente von Ctenozamites wolfiana (GOTHAN) HARRIS (Samenfarne) mit sehr gut erhaltener Kutikula werden beschrieben. Außerdem wurden mehrere Belegstücke einer Cycadophyten-Art gefunden, die wir als neue Pseudoctenis-Art (Pseudoctenis prossii) beschreiben. Leider war bei dieser Art die Kutikula an keinem einzigen Stück erhalten. Daher können wir dieses Material nicht mit Sicherheit der Gattung Pseudoctenis (Cycadeen) zuordnen; die makromorphologischen Merkmale sprechen jedoch für diese Zuteilung.
Summary
New material of Ctenozamites wolfiana (GOTHAN) HARRIS with well preserved cuticles is described while it provided additional data. Moreover, several specimens of a Cycadophyte species are described as a new species of Pseudoctenis, Pseudoctenis prossii. No cuticle was preserved in any of the specimens, and therefore, we cannot attribute the material with certainty to the genus Pseudoctenis (Cycads) but the macromorphological characters fit the genus best.
Einleitung
Obwohl die liassische Flora Bayerns mehrmals Thema einer Monographie war (SCHENK 1867, GOTHAN 1914), ist das letzte Gesamtwerk (WEBER 1968) fast 30 Jahre
alt. Seit dem sind nur Veröffentlichungen über einige separate Taxa erschienen (z.B. BOERSMA 1985, CLEMENT-WESTERHOF und VAN KONUNENBURG-VAN CITTERT 1991, VAN KONIJNENBURG - VAN CITTERT 1992,
KIRCHNER 1992, KIRCHNER & MÜLLER 1992, SCHMEISSNER & HAUPTMANN 1993, HAUPTMANN, S. & HAUPTMANN, T. 1994, KIRCHNER und VAN KONIJNENBURG-VAN CITTERT 1994).
Das ist auch nun der Fall. In einer Fundstelle bei Forkendorf/Forstmühle wurden einige kleine Fragmente von Ctenozamites wolfiana (GOTHAN)HARRIS gefunden. Nach der
Neubeschreibung GOTHANs (1914) ist dies das erste Mal, daß diese Art wieder gefunden wurde. Die Kutikula ist sehr gut erhalten, und deswegen beschreiben wir diese jetzt genauer als
GOTHAN.
Außerdem sind an verschiedenen Fundorten mehrere Belegstücke einer Cycadeen- (oder Bennettiteen-) Art gefunden worden, die wir als neue Pseudoctenis-Art beschreiben möchten, weil das Material morphologisch am besten in dieser Gattung einzu-ordnen ist. Leider war bei dieser Art die Kutikula an keinem einzigen Stück erhalten, deswegen können wir das Material nicht genauer bestimmen.
>> weiterlesen im pdf (download open access) >
- Published online 18-12-2023 by A.M. Heyng -
Eine Mikroflora aus Pechgraben und ihre stratigraphische Stellung
im Lias von Franken (S-Deutschland)
von Glenn G. Fechner
Zeitschrift Documenta naturae Bd. 117 / 1998
Kurzfassung
Proben aus einem "Rhät-Lias"-Profil der Sandgrube Dietz bei Pechgraben wurden palynologisch untersucht. Die schluffig-feinsandigen bzw. kohlig-tonigen Sedimente lieferten Pollen &
Sporen sowie aquatische Palynomorphen (Prasinophyceen, "Acritarchen", Limnoplankton und Foraminiferen-Innentapeten).
In der Mikroflora dominieren bisaccate Koniferen-Pollen. Erst mit deutlichem Abstand folgen inaperturate Pollenkörner, Cerebropollenites, Corollina und eine diverse Sporenflora.
Das marine Mikrophytoplankton setzt sich überwiegend aus den Vertretern des Micrhystridium/Veryhachium-Komplexes und einigen Formen der Gattung Tasmanites zusammen. Auf
einen gewissen Süß- bis BrackwassereinfluB deuten wenige Exemplare der Plankton-Formen Schizocystia rara und Botryococcus hin.
Anhand der Mikroflora dürfte der Untersuchungsbereich stark terrestrisch geprägt gewesen sein, jedoch mit spürbaren marinen Beeinflussungen durch gelegentliche Überflutungen, eine Fazies, wie sie
in einer Delta-Ebene zu erwarten ist. Stratigraphisch kann die Mikroflora aus Pechgraben in das mittlere bis höhere Hettang gestellt werden.
Abstract
Samples from a "Rhaetic-Liassic" sequence from a sand pit south of the village Pechgraben were examined by palynological methods. The silty to fine-sandy and coaly-clayish sediments
yielded pollen, spores and plankton (Prasinophyta, "acritarchen", limnoplankton and foraminiferal linings).
The microflora is dominated by bisaccate pollen grains. Clearly smaller is the content of inaperturate pollen grains, Cerebropollenites, Corollina and diverse spores. The marine
microphytoplankton consists beside some representatives of the genus Tasmanites mostly of the shallow water taxa Micrhystridium and Veryhachium. But a small fresh to
brackish water influence is indicated by the presence of a few specimens of Schizocystia rara and Botryococcus.
Based on the microflora the area under study is interpreted as a broad delta plain, a terrestrial environment with low or temporary marine influence.
From the stratigraphical point of view the microflora from the sand pit south of Pechgraben can be placed in the middle to upper Hettangian.
Einführung
Im Zusammenhang mit palynologisch-faziellen Untersuchungen transgressiver Abfolgen im europäischen Meso- und Käno-zoikum waren auch die Mikrofloren der Litoralfazies im fränkischen
"Rhät-Lias" von Interesse. Bei starker Vereinfachung läßt sich zwar in weiten Teilen des fränkischen "Rhät-Lias" das "terrestrische", d.h. mehr sandige Rhät vom "marinen oder transgressiven",
d.h. mehr tonigen Lias unterscheiden, doch ist im Einzelnen mit einer solchen rein lithologischen Ansprache keine verläßliche stratigraphische Zuordnung möglich. Für eine eindeutige Grenzziehung
zwischen dem Rhät und Lias haben sich in dieser Region bisher nur die Mikrofloren als brauchbar erwiesen, da sie sowohl in terrestrisehen als auch in marinen Ablagerungen, d.h.
faziesübergreifend, in ausreichender und damit auch in reproduzierbarer Menge vorkommen und so für
stratigraphisehe Untersuchungen zur Verfügung stehen.
Zwar hatte schon schon Jung (1958, 1960) über disperse Megasporen im fränkischen "Rhät-Lias" berichtet, doch sind gerade
Megasporen meist Faziesfossilien, d.h. an einen eher terrestrisehen bis litoralen Ablagerungsraum gebunden und damit für stratigraphische Aussagen nur sehr eingeschränkt nutzbar. Sierotin (1961)
legte nur wenig später eine palynologisehe Arbeit über die "Rhät-Lias"-Sedimente von Großbellhofen in Mittelfranken vor (Abb. 1), wo ein erster Versuch der palynostratigra-phischen Abgrenzen von
Rhät und Lias unternommen wurde. Eine ausführliche palynologisehe Bearbeitung rhätischer und liassischer Schichten Frankens erfolgte jedoch erst durch Achilles (1981), wo neben einigen
durchgehenden Profilen auch eine Reihe weiterer "Rhät-Lias"-Vorkommen anhand von Stichproben untersucht wurden. Da es in Franken jedoch zahlreiche Auf-
schlüsse mit "Rhät-Lias"-Ablagerungen gibt und Achilles seinerzeit nicht alle bearbeiten konnte, soll nun hier eine für die Region neue liassisehe Mikroflora aus der Lokalität "Pechgraben" (Abb.
1) vorgestellt werden.
Das Untersuchungsmaterial stammt aus der Sandgrube Dietz, die sich ca. 1 km südlich des Ortes Pechgraben befindet (TK 25 Blatt 5935, Marktschorgast; h 55 41 460, r 44 67 080). Nach Angaben von Dr. H-J. Gregor sind in der Sandgrube bis zu 18 m hohe Sedimentabfolgen aufgeschlossen. Das Liegende wird von mehreren Metern hohen grauen Sandsteinen (Arkosen?) mit eingeschalteten feineren Lagen gebildet. Darüber liegen unterschiedlich mächtige, weil lateral rasch auskeilende Schichten (u.a. graue oder rötliche Tonen mit Pflanzenfossilien, schräggeschichtete Sande, plattige Sandsteine und gelbweiße Silte). Die fünf hier für palynologisehe Untersuchungen aufbereiteten Proben hatte Dr. H.-J. Gregor aus verschiedenen grauen Ton- und hellen SiIt-(Feinsand)-lagen entnommen.
Während die zwei Feinsand-/Si1tstein-Proben keinerlei säureresistente organische Reste lieferten, enthielten die drei grauen TonProben reichlich organisches Pflanzenmaterial. In den Präparaten der Proben 4 und 5 war zwar viel Amorphogen, (feinster organischer Detritus, der im Lichtmikroskop nicht weiter differenziert werden kann; vgl. Bujak et al. 1977: 199), viel schwarzer und etwas weniger auch brauner Pflanzenhäcksel vorhanden, jedoch ließen sich keine Palynomorphen finden. Lediglich die Probe 1 lieferte eine auswertbare Mikroflora, bestehend aus Sporen, Pollen, Prasinophyceen, "Acritarchen", Limnoplankton und Foraminiferen-Innentapeten.
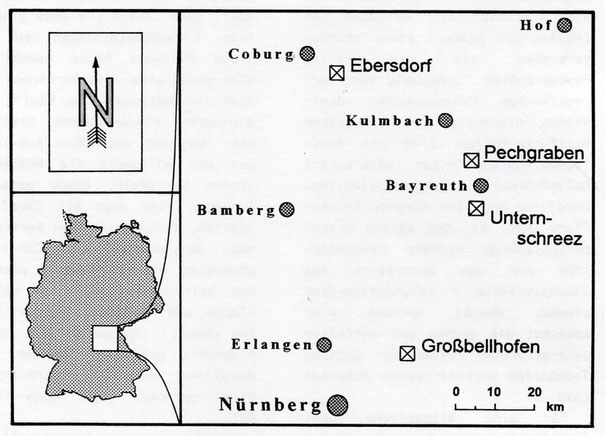
Abbildung 1. Lage der untersuchten "Rhät-Lias"-Lokalitäten in Franken.
Präparation und Dokumentation
Um das karbonatfreie Material aufzuschließen, wurden etwa 30 g
der Proben ca. 24h mit Flußsäure (38%) behandelt, (wobei die drei
Tonproben infolge der Reaktionswärme kurz aufkochten). Anschließend
konnte der säureresistente organische und anorganische Rest mit Hilfe einer salzsauren Zinkbromidlösung (d=2) und einer Zentrifuge bei 3000 min-1 getrennt werden. Für die lichtmikroskopischen Untersuchungen wurden insgesamt 18 großflächige Streupräparate in Glyzeringelatine (20 x 60mm), versiegelt mit Paraffin, angefertigt. Die Präparate sind unter den Bezeichnungen Pech 1, Pech 4 und Pech
5 in der Sammlung des Instituts für Paläontologie der Freien Universität Berlin
hinterlegt. Die bei den Abbildungserklärungen der Phototafeln angegebenen
Koordinaten beziehen sich auf das Mikroskop "Leica DMRB" beim
Landesamt für Geowissenschaften und Rohstoffe Brandenburg
(Kleinmachnow).
Fazielle und paläoklimatisehe Interpretation der Mikroflora
Die Präparate enthalten bei flüchtiger Betrachtung nur sehr
viel schwarzen und braunen Pflanzenhäcksel (u.a. Reste von Leit- oder
auch von Kutikulargeweben), relativ wenig Amorphogen (vgl. oben) und auch nur wenig Harz. Erst nach intensiver Durchmusterung der Präparate zeigte sich eine recht diverse Mikroflora aus Pollen, Sporen und marinem Phytoplankton. Quantitativ sind in der Mikroflora die terrestrisehen Palynomorphen (Pollen und Sporen) etwas stärker vertreten als das marine Phytoplankton. Innerhalb der terrestrischen Palynomorphen dominieren eindeutig die bisaccaten Koniferen-Pollen. Erst mit deutlichem Abstand folgen inaperturate Pollenkörner, Cerebropollenites, Corollina und eine diverse Sporenflora (Abb. 2). Das marine Mikrophytoplankton besteht hauptsächlich aus den Vertretern des Micrhystridium / Veryhachium-Komplexes, obwohl optisch hier zunächst die großen und auffällig gelb-gefärbten Formen der Gattung Tasmanites vorzuherrschen scheinen (Abb. 2).
Für eine klimatische bzw. fazielle Interpretation der Mikroflora muß man vorausschicken, daß (1) Pollen und Sporen nur Hinweise auf das Klima an Land liefern können. (2) Wenn Pflanzengruppen, die anhand der Pollen und Sporen identifiziert wurden, mit rezenten Taxa verglichen werden, geschieht dies meist auf dem Niveau der Gattung oder der Familie. Und (3) die ökologischen Bedürfnisse bestimmter Taxa sind in verschiedenen Regionen nicht immer gleich und können sich seit dem Mesozoikum stark verändert haben. Nun, da die rezenten ökologischen Daten nur mit Vorsicht auf das Mesozoikum zu übertragen sind, sollen die folgenden Deutungen auch nur grobe Anhaltspunkte zum damaligen Klima bzw. Environment liefern.
Zwar sind die von verschiedenen Koniferengruppen stammenden bisaccaten Pollenkörner hier am häufigsten, doch lassen sie kaum eindeutige Klimabeurteilungen zu, da diese Pflanzen heute sowohl in kühl-gemäßigten Klimabereichen als auch in Montangebieten häufig das dominante Florenelement stellen, aber durchaus auch die (Sub-)Tropen und allgemein die Küstenregionen besiedeln. Etwas genauere Aussagen sind aber mit Corollina möglich, einem typischen Vertreter des Meso-zoikums: "...Corollina producing plants probably preferred well-drained soils of upland slopes and lowlands not far from the coast." (Srivastava 1976: 454). Allgemein soll eine Dominanz von Corollina für ein warm-arides Klima sprechen (vgl. Alvin 1982: 87).

Abbildung 2. Liste der in der Lokalität Pechgraben angetroffenen Palynomorphen.
Obwohl der Name Taxodiapollenites hiatus eine Verwandtschaft
zur rezenten Gymnospermenfamilie der Taxodiaceae suggerieren könnte, ist sie jedoch nicht gesichert (Taf. 2, Abb. 4). Zwar wurden von diesen sehr
charakteristischen inapertu-raten Pollenkörnern hier nur vereinzelte Exemplare
angetroffen, doch sind sie trotzdem faziell interessant. Ihr
Auftreten in marinen Schichten könnte bei aller gebotenen Vorsicht auf
einen möglichen "Taxodiaceae-Cupressaceae-Sumpfwald" nahe der Küste
hinweisen.
Für wahrscheinlich etwas feuchtere Verhältnisse sprechen die inaperturaten Pollenkörner, die u.a. von Araucaria-artigen Koniferen stammen dürften. Ähnliches kann auch für die monosulcaten Pollenkörner gelten, die von Cycadophyten, Ginkgo-phyten und verwandten Gymnospermen gebildet werden, Pflanzen also, die heute eher in den feuchten Tropen bis warmge-mäßigten Breiten beheimatet sind. Allerdings sind monosulcate Pollenkörner im Verhältnis zu den Makrofunden der monosulcate Pollenkörner-produzierenden Pflanzen meist deutlich unterrepräsentiert ( Frederiksen 1980: 1).
Die für mesozoische Ablagerungen typischen kleinen bisaccaten Pollenkörnen von Vitreisporites pallidus (Taf. 2, Abb. 6-7) stammen mit einiger Sicherheit von Pflanzen der Caytoniales, zumindest wurden sie zusammen mit fertilen Organen von Caytonia gefunden (vgl. Harris 1951). Diese oder vergleichbare Pteridospermen dürften eher doch feuchtere Lebensräume bevorzugt haben.
Auch mit Hilfe des Planktons sind einige Aussagen zur
Fazies möglich. Das Verhältnis marines/nichtmarines Plankton ist
durch ein eindeutiges Übergewicht der marinen Taxa gekennzeichnet.
Zwar könnte die Süßwasser-Planktonform Schizocystia rara (Taf. 2,
Abb. 13) auf lakustrine Ablagerungen hinweisen, was übrigens in guter
Übereinstimmung mit der Dominanz bisaccater und inaperturaten Pollenkörnern in der Mikroflora steht, die aufgrund ihrer hydrodynamischen Eigenschaften ja auch für solche Ablagerungen sprechen können, doch ist hier von Schizocystia rara nur ein einziges Exemplar vorhanden, so daß Seesedimente eher unwahrscheinlich sind. Auch von der im Süß- und Brackwasser vorkommenden Plankton-Form Botryococcus (Taf. 2, Abb. 12) wurde nur ein Exemplar angetroffen. Dagegen sind innerhalb der marinen Plankton-Taxa allein
von Micrhystridium (Taf. 2, Abb. 20) über 100 und von
Tasmanites (Taf. 2, Abb. 14-15) über 50 Exemplare beobachtet worden.
Die anderen marinen Plankton-Formen Cymatiosphaera,
Pleurozonaria?, Pterosphaeridia (Taf. 2, Abb. 17-19) und
Veryhachium reductum treten dagegen nur vereinzelt auf. Während
Micrhystridium (Wall & Dale 1970: 48) und Tasmanites überwiegend Flachwasserformen sind, kann die unverwechselbare Plankton-Art Veryhachium reductum (Taf. 2, Abb. 21), als ein recht zuverlässiger Warmwasser-Indikator gelten (vgl. Fechner 1989: 61), der möglicherweise auch auf eine verringerte Salinität hinweist.
Zwar sind hier nur sehr wenige rotaliide Foraminiferen-Innentapeten angetroffen worden, doch sind die derben organischen Innenhüllen von überwiegend benthonischen Foraminiferen (Taf. 2, Abb. 16) weitere sichere Indikatoren für einen marinen Einfluß.
Anhand der Pollen & Sporen kann man hier für das Festland im Lias ein eher feucht-warmes und entlang der Küstenlinie vielleicht ein etwas trockneres Klima annehmen. Das vorrückende Meer war gekennzeichnet durch flachmarine Warmwasser-Verhältisse mit vorübergehend, möglicherweise tidal-bedingten verringerten Salinitäten. Insgesamt sprechen die Daten für Sedimentationsverhältnisse, wie sie typischerweise in Deltaebenen anzutreffen sind, die gelegentlich, bei größeren Stürmen, kurzfristig vom Meer überfluten werden.
Stratigraphische Einstufung der Mikroflora
Das Alter der Mikroflora aus Pechgraben läßt sich relativ
einfach durch Kombination der unterschiedlichen stratigraphisehen
Verbreitungsdaten einiger weniger Palynomorphen ermitteln. So ist
Ricciisporites tuberculatus (Taf. 1, Abb. 22-23) vom tiefen Rhät
bis zum höchsten Hettang und Lycopodiacidites rugulatus (Taf. 1,
Abb. 21) von der Basis des Hettang bis zum höchsten Pliensbach zu
finden (vgl . Morbey & Dunay 1978: 50-51 ), weshalb für das Alter der
hier untersuchten Mikroflora nur das Hettang an Frage kommt. Da
Porcellispora longdonensis, eine große alete und echinate Spore im
Untersuchungsmaterial nicht nur recht auffällig ist, sondern auch
relativ häufig auftritt, soll deren stratigraphisehe Verbreitung hier nun auch noch betrachtet werden (Taf. 1, Abb. 16-20). Verschiedene Autoren geben für die stratigraphisehe Verbreitung von P. longdonensis in Europa nur Muschelkalk bis Rhät an (Van Erve 1977: 67). Auch nach Morbey (1975: 24 + Abb. 25) und Lund (1977: 57) soll P. longdonensis in den Alpen bzw. im Nordsee-Becken nur bis ins höchste Rhät reichen. Diese Verbreitungsangaben dürften wohl aber lediglich Folge lokaler Faziesbedingungen sein, eine Ansicht, die übrigens schon Van Erve (1977: 28) geäußert hatte, da er P. longdonensis an verschiedenen Lokalitäten des alpinen Lias nachweisen konnte. Auch für den fränki-schen Lias alpha 1 + 2 (=
Hettang) ist P. 1ongdonensis bereits durch Achilles (1981: 41) sicher belegt
worden, wodurch die oben angegebene stratigraphische Einstufung der Mikroflora
aus Pechgraben bestätigt wird.
Da das angetroffene Mikroplankton fast ausschließlich
aus Faziesfossilien besteht, kann es zur stratigraphisehen Einstufung
leider nur wenig beisteuern. Lediglich Schizocystia rara soll
hier genannt werden. Diese offenbar reine Süßwasserform war mit
großer Wahrscheinlichkeit bereits im "Rhaet-Lias" ein Kosmopolit,
da sie einerseits in terrestrischen Ablagerungen Australiens gefunden
wurde (Playfort & Dettmann, 1965: 160), andererseits Schulz & Mai
(1966b) eine solche Form in ihrer "Tabelle der Verteilung des Phytoplanktons im Lias und Dogger" als "Plankton 1" abbildeten und betonten, daß diese Form erst ab dem Lias alpha auftritt (ibid. : 41). Auch Lund (1977: Tafel 12) erwähnt diese Plankton-Form aus dem Munkerup-Clay (Munkerup-Member = Unter-Hettang) von Bornholm.
Da hier keine Vertreter von Densoisporites, Zebrasporites interscriptrus, Kraeuselisporites reissingeri oder Ovalipollis ovalis beobachtet worden sind, Formen, die sowohl im Rhät als auch im Hettang vorkommen können (vgl. Morbey & Dunay 1978: 50-51), ist ein rhätisches Alter der Mikroflora aus Pechgraben sicher auszuschließen und auch das tiefste Hettang ist nicht sehr wahrscheinlich. Damit steht die stratigraphische Einstufung der Mikroflora aus Pechgraben fest, d.h. mittleres bis höheres Hettang.
Vergleiche mit anderen Mikrofloren aus dem fränkischen "Rhät-Lias"
Zwar kann man zur regionalen Beurteilung der Mikroflora aus
Pechgraben auf einige palynologische Literaturdaten zum "Rhät-Lias"
von Franken zurückgreifen, doch ist "eigenes" Vergleichsmaterial in
der Regel besser geeignet, besonders, weil unter-schiedliche
Aufbereitungsmethoden der Gesteinsproben einen nicht unerheblichen
Einfluß auf die Zusammensetzung der Mikrofloren haben können. Für
Vergleichsuntersuchungen wurden einige Proben aus den
"Rhät-Lias"-Lokalitäten Großbell-hofen, Unternschreez und Ebersdorf
verwendet (Abb. 1). Während in der Tongrube bei Ebersdorf bereits ein
vollständiges Profil palynologisch bearbeitet wurde (Achilles 1981),
sind aus der Sandgrube Unternschreez lediglich wenige, im Einzelnen nicht genauer dokumentierte, "Stichproben" untersucht worden (ibid.). Auch aus den zwei Gruben bei Großbellhofen hat Sierotin (1961) lediglich fünf nicht zusammenhängende Proben bearbeitet. Das hier verwendete Vergleichsmaterial aus Großbellhofen wurden mir freundlicherweise von Dr. H-J. Gregor zur Verfügung gestellt; das Material aus Unternschreez und Ebersdorf stammt aus eigenen Beprobungen.
Großbellhofen in Mittelfranken, östlich von Nürnberg bei Schnaittach, liegt ca. 50 km südlich von Pechgraben (Abb. 1), wo nach Angaben von Sierotin (1961: 7) in zwei Gruben (große und blaue Grube) Ton gewonnen wurde. Auch die Lokalität "Wolfshöhe", wo Achilles (1981: 68) einige Stichproben gesammelt hatte, befindet sich nahe der Ortschaft Schnaittach. Die hier untersuchte Tonprobe aus Großbellhofen lieferte reichlich organisches Material (mäßig-viel Amorphogen, viel schwarzer und brauner Pflanzenhäcksel, sehr wenig Harz) und eine diverse Mikroflora. Zwar dominieren in der Mikroflora bisaccate Pollenkörner, doch sind auch Corollina (z.T. als Tetraden) und Vitreisporites pallidus ausgesprochen häufig anzutreffen. Cycadopites und Inaperturopollenites sind dagegen nur mäßig-häufig und T. hiatus ist sogar recht selten. Von der relativ diversen Sporenflora sollen nur wenige charakteristische Formen genannt werden (häufig Camarozonosporites (z.T. auch als Tetraden), selten dagegen Striatella sp. und Deltoidospora toralis). Da typische Hettangformen, wie in Pechgraben (u.a. Poicellispora longdonensis, Ricciisporites tuberculatus, PIatysaccipollenites, Lycopodiacidites rugulatus) fehlen, dürfte die Vergleichsprobe deutlich jünger sein als das Material aus Pechgraben, zumal auch marine Anzeiger fehlen.
Überraschenderweise zeigt die hier untersuchte Vergleichsprobe aus Großbellhofen die größten Übereinstimmungen mit den Proben, die von Sierotin (1961: 57) ins "Rhät" gestellt wurden (d.h. viel Corollina und viel Vitreisporites pallidus), eine strati-graphisehe Einstufung, die, da Sierotin keine obligat-rhätisehen Elemente genannt bzw. abgebildet hat und auch hier keine nachgewiesen wurden, wohl recht zweifelhaft ist. Marine Hinweise sind bei Sierotin (1961: 57) zwar im "Lias alpha" und "Lias gamma" "Acanthotriletes bellhofensis" bzw. im "Lias gamma" "Foliopollenites spinosus" zu finden, (beide Formen sind wohl zu Micrhystridium zu rechnen), doch sind all diese stratigraphisehen Einschätzungen recht unsicher, zumal dort auch keinerlei echten "Rhät-Lias"-Formen zu finden sind. Entweder sind die Ablagerungen bei Großbellhofen deutlich jünger als Hettang oder aber es handelt sich um eine völlig ungewöhnliche, wohl nichtmarine Fazies (vgl. auch Achilles 1981: 11; "Wolfshöhe"). Zur Klärung dieser Fragen sollten dort einige Profile genauer untersucht werden.
Unternschreez, unweit Bayreuth, liegt nur ca. 15 km südlich von Pechgraben entfernt (Abb. 1). In der Sandgrube befinden sich hier oberhalb des hellen "Pflanzensandsteins" geringmächtige, lateral oft rasch auskeilende Lagen von (Fein-)Sandsteinen und tonigen Partien, die z.T. zahlreiche Pflanzenfossilien und gelegentlich auch Rhizocorallium enthalten, was auf eine Litoral-fazies hinweist. Eine in diesem Bereich entnommene Probe, ein "kohliger Kaolinit" (Pr.-Nr. 8722), lieferte reichlich organisches Material (wenig Amorphogen, viel schwarzer aber nur mäßig-viel brauner Häcksel, sehr wenig Harz) und eine auswertbare Mikroflora. Während bisaccate Pollen und Sporen (z.T. als Trauben) regelmäßig vorhanden sind und auch Cerebropollenite nicht selten ist, sind jedoch nur relativ wenige Exemplare von Corollina (z.T. auch als Tetraden) und Cycadopites beobachtet worden. Inaperturopollenites sowie T. hiatus treten sogar nur vereinzelt auf.
Obwohl die Ablagerungen von Unternschreez mit denen von Pechgraben lithofaziell recht gut vergleichbar sind, wurden hier jedoch keine der typischen Formen wie Vitrei sporites pallidus, Porcellispora longdonensis, Ricciisporites tuberculatus, PIatysaccipoliertites, Lycopodiacidites rugulatus und auch kein marines Plankton gefunden. Entweder ist der Lias von Unternschreez jünger als der von Pechgraben oder es handelt sich eine ungewöhnliche Litoralfazies. Zwar ist bei Achilles (1981: 65) die Lokalität Unternschreez nicht ausdrücklich genannt, doch rechnete er die Mikrofloren des "Bayreuther Gebietes" zu einem "wahrscheinlichen" Lias alpha 2 (= höheres Hettang).
Ebersdorf, östlich von Coburg, liegt ca. 40 km nordwestlich von Pechgraben (Abb. 1). Die Firma "ESTO-Klinker" baut dort in einem ausgedehnten Grubenkomplex Tone des "Rhät-Lias" ab. Da eine ausführliche Profil-Untersuchung bereits durch Achilles (1981: 9-10) erfolgte, wurde hier nur eine Probe aus dem untersten Hettang zum Vergleich herangezogen. Die Probe (Pr.-Nr. 8718) stammt aus einem mittel grauen feinblättrigen Tonstein, ca. 120 cm unterhalb des am Probennahme-Punkt mindestens 90 cm mächtigen "Ebersdorfer Sandsteins". Danach gehört das Material in den Lias alpha 1 (= tieferes Hettang; Psiloceras-Schichten).
Die Probe lieferte ausreichend organisches Material (wenig
Amorphogen, mäßig-viel schwarzer und nur wenig brauner Häcksel, sehr
wenig Harz, aber sehr viele Pellets) und eine relativ arme Mikroflora. Die bisaccaten Pollenkörner, Corollina (z.T. Tetraden + Trauben) und Inaperturopollenites sind überraschenderweise jeweils nur mit wenigen Exemplaren angetroffen worden. Dagegen ist der Anteil der Sporen (z.T. als Tetraden oder Trauben) relativ groß. Von den für die stratigraphische
Beurteilung wichtigen Formen sind Porcellispora
longdonensis, Ricciisporites tuberculatus und Lycopodiacidites
rugulatus jeweils mit einzelnen, Kraeselisporites sogar mit mehreren
Exemplaren vorhanden. Einige Vertreter der Gattung Tasmanites (Durchmesser
ca. 55-70 Mikrometer) sind ein deutlicher Hinweis auf eine marine Beeinflussung
der Abiagerungen.
Wenn man von Kraeselisporites absieht, eine Form, die in Pechgraben nicht nachzuweisen war, ist die Mikroflora aus dem tiefsten Hettang von Ebersdorf stratigraphisch und faziell der Mikroflora aus Pechgraben am ähnlichsten. Interessant dabei ist jedoch, daß sich gerade aufgrund der Abwesenheit von Kraeselisporites in der Mikroflora von Pechgraben, dort das tiefste Hettang wahrscheinlich ausschließen läßt, womit die bereits oben erfolgte stratigraphische Einstufung der Mikroflora aus Pechgraben ins mittlere bis höhere Hettang bestätigt wird.
Literatur
Achilles, H. (1981): Die rätische und liassische
Mikroflora Frankens. - Palaeontographica, 179(1-4): 1-86, 32
Abb., 1 Tab., 17 Taf.; Stuttgart.
Alvin, K.L. (1982): Cheirolepidiceae: Biology, structure
and paleoecology. - Review of Palaeobotany and
Palynololgy, 37: 71-98, Taf. 1, 12 Abb.; Amsterdam.
Bujak, J.P., Barss, M.S. & Williams, G.L. (1977): Offshore
East Canada's organic type and color hydrocarbon potential
- Part I. - The Oil and Gas Journal, 1977(4): 198-202, 4 Abb.; Tulsa, Oklahoma.
Clarke, R.F.A. (1965): Keuper miospores from
Worcestershire, England. - Palaeontology, 8(2): 294-321, 13
Abb., Taf. 35-39; London.
Cookson, I.C. (1947): Plant microfossils from the lignites
of Kerguelen Archipelago. British-, Australian-, New
Zealand Antarctic Research Expedition (1929-1931) [B.A.N.Z.A.R.E. ], Science Reports, Series A, Vol. II, Part 8:
127-142, Taf. 13-17.
Couper, R.A. (1953): Upper Mesozoic and Cainozoic spores
and pollen grains from New Zealand. - New Zealand
Department of Scientific and Industrial Research,
Paleontological Bulletin, 22: 1-77, Taf. 1-9; Wellington.
Couper, R.A. (1958): British mosozoic microspores and
pollen grains, systematic and Stratigraphie study. -
Palaeontographica, B 103(4-6): 75-179, 11 Abb., 12 Tab., Taf.
15-31; Stuttgart.
Danze-Corsin, P. & Laveine, J.-P. (1963): Microflore. In:
Eriche, P., Danze-Corsin, P. & Laveine, J.-P.: Flore
infraliasique du Boulonnais (Macro- et Microflore).
Societe Geologique du Nord, Memoires, 13: 57-143, Abb. 5, Tab. A-D, Taf. 5-11; Lille.
Deflandre, G. (1935): Microorganismes d’origine
planctonique conserves dans les silex de la Craie. - Bulletin
biologique de la France et de la Belgique, 69(2): 213-244,
Taf. 5-9; Paris.
Deflandre, G. (1937): Microfossilles des silex Cretaces
III-VI, Flagelles incertae sedis. Hystrichosphaerides.
Sarcodines Organisms divers. Annales de Paläontologie,
26: 51-103, Taf. 8-15, Paris.
Deunff, J. (1958): Micro-organismes planctoniques du
Primaire armoricain. 1. Ordovicien du Veryhach (Presqu'ile de
Crozon). - Bulletin de la Societe geologique et mineralogique de Bretagne, N.S., 2: 1-41, 12 Taf.; Rennes.
Dybkj/er, K. (1991): Palynological zonation and
palynofacies Investigation of the Fjerritslev Formation (Lower
Jurassic - basal Middle Jurassic) in the Danish Subbasin.
- Danmarks Geologiske Underspgelse, Serie A, Nr. 30: 1-150, 21 Abb., 22 Taf., Appendix A-F; Kpbenhavn.
Fechner, G.G. (1989): Palynologisehe Untersuchungen im
Alb/Cenoman-Grenzbereich von Rüthen (NW-Deutschland) und
La Vierre (SE-Frankreich). - Documenta naturae, 53: 1—136 + i-xiii Taf. 1-34; München.
Frederiksen, N.O. (1980): Significance of monosulcate
pollen abundance in Mesozoic Sediments. - Lethaia, 13: 1-20;
Oslo.
Harris, T.M. (1951): The fructification of Czekanowskia
and its al lies. - Philosophical Transactions of the Royal
Society of London, B 235: 483-508; London.
Jekhowsky, de B. (1961): Sur quelques hystrichospheres
Permo-Triassiques d'Europe et d'Afrique. - Revue de
Micropaleontologie, 3(4): 207-212, Taf. 1-2; Paris.
Jung, W. (1958): Zur Biologie und Morphologie einiger
disperser Megasporen, vergleichbar mit solchen von
Lycostrobus
scotti, aus dem Rhät-Lias Frankens. -
Geologische Blätter für Nordost-Bayern und angrenzende Gebiete, 8:
114-130, 2 Abb., Taf. 3; Erlangen.
Jung, W. (1960): Die dispersen Megasporen der
fränkischen Rhät-Lias-Grenzschichten. Palaeontographica, B
107(4-6): 127-170, 1 Tab., Taf. 36-39; Stuttgart.
Klaus, W. (1960): Sporen der karnischen Stufe der
ostalpinen Trias. - Geologisches Jahr buch der Bundesanstalt Wien,
Sonderband, 5: 107-184, Taf. 28-38; Wien.
Kremp, G. (1949): Pollenanalytische Untersucungen des
Miozänen Braunkohlenlagers von Konin an der Warthe. -
Palaeonto-
graphica, B 90: 53-93, 18 Abb., Taf. 3-9;
Stuttgart.
Leschik, G. (1955): Die Keuperflora von Neuewelt bei Basel.
II. Die Iso- und Mikrosporen. - Schweizerische
Palaeontolo-
gische Abhandlungen, 72: 1-70, Taf. 1-10;
Basel.
Lund, J.J. (1977): Rhaetic to Lower Liassic palynology of
the onshore south-eastern North Sea Basin. - Danmarks
Geolo-
giske Underspgelse,II Raekke, Nr. 109: 1-129, 5 Abb., 4
Tab., 12 Taf., Appendix A-F; Kobenhavn.
Lundblad, B. (1954): Contributions to the Geological History
of the Hepaticae. Fossil Marchantiales from the
Rhaetic-Liassic coal mines of Skromberga (Prov. of Scania),
Sweden. - Svensk Botanisk Tidskrift, 48(2): 381-417; Stockholm.
Mädler, K. (1963): Die figurierten organischen Bestandteile
der Posidonienschiefer. - Beihefte zum Geologischen
Jahrbuch, 58: 287-406, 4 Abb., 3 Tab., 16 Taf.;
Hannover.
Mädler, K. (1964): Bemerkenswerte Sporenformen aus dem
Keuper und unteren Lias. - Fortschritte in der Geologie von
Rheinland und Westfalen, 12: 169-200, 3 Taf., 1 Tab.; Krefeld.
Morbey, S.J. (1975): The palynostratigraphy of the
Rhaetic stage, Upper Triassic in the Kendelbachgraben, Austria. -
Palaeontographica B 152 (1-3): 1-75, 37 Abb., 3 Tab., 19 Taf.; Stuttgart.
Morbey, S.J. & Dunay, R.E. (1978): Early Jurassic to Late
Triassic dinoflagellate cysts and Miospores. In: Thusu, B.
[Hrsg.]: Distribution of biostratigraphically diagnostic dinoflagellate cysts and miospores from the Northwest
European Continental
shelf and adjacent areas. - IKU, Institutt for
Kontinentalsokkelunderspkelser, No. 100: 47-59, 1 Tab., 4 Taf.;
Trondheim, Norwegen.
Nilson, T. (1958): Über das Vorkommen eines mesozoischen
Sapropelgesteins in Schonen. - Lunds Universitets
Ärsskrift, N.F. Avd. 2, Band 54(10): 1-111, Taf. 1-8;
Lund.
Playford, G. & Dettmann, M.E. (1965): Rhaeto-Liassic plant
microfossils from the Leigh Creek coal measures, South
Australia. - Senckenbergia Lethea, 46(2/3): 127-181,
Taf. 12-17, 1 Abb.; Frankfurt.
Potonie, R. (1932): Pollenformen aus tertiären Braunkohlen.
- Jahrbuch der preußischen geologischen Landesanstalt
zu Berlin für das Jahr 1931, Band 52: 1-7, 34 Abb. Berlin.
Potonie, R. (1958): Synopsis der Gattungen der Sporae
dispersae. II. Teil: Sporites (Nachträge),
Saccites, Aletes, Praecolpates, Polyplicates, Monocolpates. -
Beihefte zum Geologischen Jahrbuch, 31: 1-114, Taf. 1-11;
Hannover.
Potonie, R. & Klaus, W. (1954): Einige Sporengattungen des
alpinen Salzgebirges. - Geologisches Jahrbuch, 68:
517-544, 11 Abb., Taf. 10; Hannover.
Reissinger, A. (1950): Die "Pollenanallyse" ausgedehnt auf
alle Sedimentgesteine der geologischen Vergangenheit.
Palaeontographica B 90: 99-126, Taf. 11-19; Stuttgart.
Scheuring, B.W. (1970): Palynologische und
palynostratigraphische Untersuchungen des Keuper im Bölchentunnel
(Solo-
thurner Jura). - Schweizerische Pal aeontologisehe
Abhandlungen, 88: 1-119, 5 Abb., 57 Tab., 43 Taf.;
Basel.
Schulz, E. (1967): Sporenpaläontologische Untersuchungen
rätoliassischer Schichten im Zentralteil des Germanischen
Beckens. - Paläontologische Abhandlungen Abt. B, Paläobotanik, Band II, Heft 3: 541-633, 26 Taf., 2 Abb, 1 Tab.;
Berlin.
Schulz, E. & Mai, D.H. (1966b): Erläuterungen zur Tabelle
der stratigraphisehen Verbreitung des Phytoplanktons im
Lias und Dogger. In: Döring, H., Krutzsch , W. , Mai , D.H. &
Schulz, E. [Hrsg.]: Erläuterungen zu den sporenstratigraphischen Tabellen vom Zechstein bis zum Oligozän. -
Abhandlungen des Zentralen Geologischen Instituts, Heft 8: 35-45, 1 Tab.;
Berlin.
Sierotin, T. (1961): Sporae dispersae im Rhät und Lias
von Grossbellhofen (Mittelfranken). - Unpublizierte
Dissertation an der Freien Universität Berlin: 1-77, 5
Taf.; Berlin.
Srivastava, S.K. (1976): The fossil pollen genus Classopolis.
- Lethaia, 9: 437-457; Oslo.
Van Erve, A.W. (1977): Palynological Investigation in the
Lower Jurassic of the Vicentinian Alps (Northeastern Italy). -
Review of Palaeobotany and Palynology, 23: 1-117, 9 Abb., 22 Taf.; Amsterdam.
Wall, D. & Dale, B. (1970): Living hystrichosphaerid
dinoflagen ate spores from Bermuda and Puerto Rico. -
Micropaleontology, 16(1): 47-58, Taf. 1; New York.
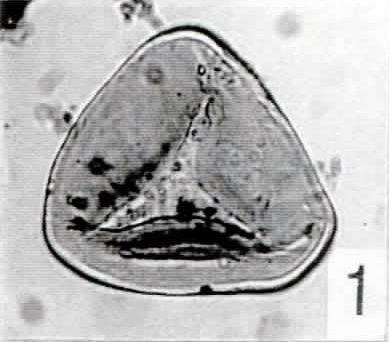
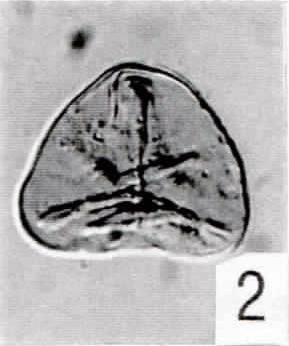
Tafel 1
(Alle Abbildungen x500; IC = Interferenzkontrast)
1. Cyathidites australis (Pech 1-02: 42,1/102,5)
2. Cyathidites minor (Pech 1-01: 53,3/98,3) (IC)
3. cf. Uvaesporites (Pech 1-13: 34,5/105,4)
4. cf. Leptolepidites [Pech 1-02: 30,2/101,1]
5. Camarozonosporites rudis [Pech 1-09: 48,7/107,8]
6. Striatella jurassica [Pech 1-03: 36,7/100,2]
7. Striatella sp. [Pech 1-05: 68,3/109,5]
8. Trachysporites sp. [Pech 1-12: 68,1/106,6] (IC)
9. Spore "A" [Pech 1-12: 31,0/97,6]
10. Deltoidospora toralis [Pech 1-12: 44,2/101,7]
11. Leiotriletes sp. [Pech 1-03: 48,5/95,0] (IC)
12. Osmundacidites aff. wellmannii [Pech 1-02: 61,6/106,7] (IC)
13. ?Leptolepidites-Tetrade [Pech 1-05: 67,6/102,0]
14. Spore "B" [Pech 1-02: 41,3/101,4] (IC)
15. Spore "C" [Pech 1-10: 48,7/102,6] (IC)
16. - 17. Porcellispora longdonensis [Pech 1-01: 46,4/99,7]
18. - 19. Porcellispora longdonensis [Pech 1-06: 32,7/103,0]
20. Porcellispora longdonensis [Pech 1-01: 31,0/104,3]
21. Lycopodiacidites rugulatus [Pech 1-05: 29,0/111,6]
22. Ricciisporites tuberculatus [Pech 1-07: 35,9/114,6]
23. Ricciisporites tuberculatus [Pech 1-08: 62,1/112,8]
click to enlarge & read figure captions
![4. cf. Leptolepidites [Pech 1-02: 30,2/101,1]](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/id6b9e7eb69ce72c9/version/1690722291/image.jpg)
![5. Camarozonosporites rudis [Pech 1-09: 48,7/107,8]](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i78f8be965c057890/version/1690722291/image.jpg)
![6. Striatella jurassica [Pech 1-03: 36,7/100,2]](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i6815f6cf1e39e3cb/version/1690722291/image.jpg)
![7. Striatella sp. [Pech 1-05: 68,3/109,5]](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i0480f4028b4766bd/version/1690722291/image.jpg)
![8. Trachysporites sp. [Pech 1-12: 68,1/106,6] (IC)](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i11457ed58fb5c813/version/1690722291/image.jpg)
![9. Spore "A" [Pech 1-12: 31,0/97,6]](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/ib18c94856ef9de76/version/1690722291/image.jpg)
![10. Deltoidospora toralis [Pech 1-12: 44,2/101,7]](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i76f31c74528be550/version/1690722291/image.jpg)
![11. Leiotriletes sp. [Pech 1-03: 48,5/95,0] (IC)](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/ifed2a9892ba05847/version/1690722291/image.jpg)
![12. Osmundacidites aff. wellmannii [Pech 1-02: 61,6/106,7] (IC)](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/iac4fa4c823521148/version/1690722291/image.jpg)
![13. ?Leptolepidites-Tetrade [Pech 1-05: 67,6/102,0]](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i6b0bf81057ca0f75/version/1690722291/image.jpg)
![14. Spore "B" [Pech 1-02: 41,3/101,4] (IC)](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/if0a3305dea98fb0e/version/1690722291/image.jpg)
![15. Spore "C" [Pech 1-10: 48,7/102,6] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i3486dbefc00ed1cf/version/1690722291/image.jpg)
![16. Porcellispora longdonensis [Pech 1-01: 46,4/99,7]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i786f59302ee52072/version/1690722291/image.jpg)
![17. Porcellispora longdonensis [Pech 1-01: 46,4/99,7]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/ie57fac8a7cf2f05e/version/1690722291/image.jpg)
![18. Porcellispora longdonensis [Pech 1-06: 32,7/103,0]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/ic3e78525685309b7/version/1690722291/image.jpg)
![19. Porcellispora longdonensis [Pech 1-06: 32,7/103,0]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i1ea75acb334e12a2/version/1690722291/image.jpg)
![20. Porcellispora longdonensis [Pech 1-01: 31,0/104,3]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/ia60b745064432d94/version/1690722291/image.jpg)
![21. Lycopodiacidites rugulatus [Pech 1-05: 29,0/111,6]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i6320666b5b041e13/version/1690722291/image.jpg)
![22. Ricciisporites tuberculatus [Pech 1-07: 35,9/114,6]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i44438e7a24c8097b/version/1690722291/image.jpg)
![23. Ricciisporites tuberculatus [Pech 1-08: 62,1/112,8]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i74a02c64be45190c/version/1690722291/image.jpg)
Tafel 2
(Abbildungen 1-19 x500, Abbildungen 20-21 x1000; IC = Interferenzkontrast)
1. Platysaccipollenites cf. papilionis [Pech 1-14: 59,4/106,3] (IC)
2. "Alisporites" sp. [Pech 1-01: 50,4/101,0]
3. bisaccates Pollenkorn indet. [Pech 1-03: 59,5/100,6] (IC)
4. Taxodiaceaepollenites hiatus [Pech 1-09: 34,8/102,7] (IC)
5. Monosulcites minimus [Pech 1-04: 34,7/95,5] (IC)
6. Vitreisporites pallidus [Pech 1-01: 24,2/104,4] (IC)
7. Vitreisporites pallidus [Pech 1-02: 60,7/106,6] (IC)
8. Cerebropollenites mesozoicus [Pech 1-04: 48,7/106,3] (IC)
9. Inaperturopollenites sp. [Pech 1-14: 54,7/107,4] (IC)
10. "Spheripollenites psilatus Couper 1958" sensu Dybkjaer 1991 (Taf. 13, Fig 7) [Pech 1-11: 35,2/99,5]
11. Corollina-Tetrade [Pech 1-10: 40,2/98,4]
12. Botryococcus sp. [Pech 1-03: 29,0/111,1] (IC)
13. Schizocystia rara [Pech 1-08: 20,6/98,7] (IC)
14. Tasmanites sp. 1 [Pech 1-09: 30,5/106,3] (IC)
15. Tasmanites sp. 2 [Pech 1-01: 32,8/101,5]
16. rotaliide Foraminiferen-Innentapete [Pech 1-05: 65,2/101,4] (IC)
17. - 18. Pterosphaeridia nodosa [Pech 1-12: 18,7/101,0] (IC)
19. Pterosphaeridia sp. [Pech 1-14: 44,0/111,5]
20. Micrhystridium sp. [Pech 1-02: 47,3/106,6] (IC)
21. Veryhachium reductum [Pech 1-07: 31,6/94,6] (IC)
click to enlarge & read figure captions
![1. Platysaccipollenites cf. papilionis [Pech 1-14: 59,4/106,3] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i3ede41ce0c46c14f/version/1690721356/image.jpg)
![2. "Alisporites" sp. [Pech 1-01: 50,4/101,0]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i8ad9aa5ab6515d8b/version/1690721357/image.jpg)
![3. bisaccates Pollenkorn indet. [Pech 1-03: 59,5/100,6] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i8edf10250569825d/version/1690721357/image.jpg)
![4. Taxodiaceaepollenites hiatus [Pech 1-09: 34,8/102,7] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/idb8b5ad26e58eb28/version/1690721357/image.jpg)
![5. Monosulcites minimus [Pech 1-04: 34,7/95,5] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i9030680b5ec64126/version/1690721357/image.jpg)
![6. Vitreisporites pallidus [Pech 1-01: 24,2/104,4] (IC)](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i2c51a94953557f23/version/1690721357/image.jpg)
![7. Vitreisporites pallidus [Pech 1-02: 60,7/106,6] (IC)](https://image.jimcdn.com/app/cms/image/transf/none/path/s602cffe28acf71e3/image/i673fdf5b17908564/version/1690721357/image.jpg)
![8. Cerebropollenites mesozoicus [Pech 1-04: 48,7/106,3] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i4985b2b543d0bfb2/version/1690721357/image.jpg)
![9. Inaperturopollenites sp. [Pech 1-14: 54,7/107,4] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i2bf0b3b0d0e3a479/version/1690721357/image.jpg)
![10. "Spheripollenites psilatus Couper 1958" sensu Dybkjaer 1991 (Taf. 13, Fig 7) [Pech 1-11: 35,2/99,5]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/ib6c93d63ff6bd4c8/version/1690721357/image.jpg)
![11. Corollina-Tetrade [Pech 1-10: 40,2/98,4]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i4301d199c60145a3/version/1690721357/image.jpg)
![12. Botryococcus sp. [Pech 1-03: 29,0/111,1] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i38080ac9b01e994d/version/1690721357/image.jpg)
![13. Schizocystia rara [Pech 1-08: 20,6/98,7] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/id0c5b57ac57859df/version/1690721357/image.jpg)
![14. Tasmanites sp. 1 [Pech 1-09: 30,5/106,3] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i7471babeab68a910/version/1690721357/image.jpg)
![15. Tasmanites sp. 2 [Pech 1-01: 32,8/101,5]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i398b0745c6fca4bd/version/1690721357/image.jpg)
![16. rotaliide Foraminiferen-Innentapete [Pech 1-05: 65,2/101,4] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i14f3dd4cd5421d18/version/1690721357/image.jpg)
![17. Pterosphaeridia nodosa [Pech 1-12: 18,7/101,0] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i1e9dec90c1a68a0a/version/1690721357/image.jpg)
![18. Pterosphaeridia nodosa [Pech 1-12: 18,7/101,0] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i02a85336629f33eb/version/1690721357/image.jpg)
![19. Pterosphaeridia sp. [Pech 1-14: 44,0/111,5]](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i9f1011343e8ea1bb/version/1690721357/image.jpg)
![20. Micrhystridium sp. [Pech 1-02: 47,3/106,6] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/iae905f8dcfa20e97/version/1690721357/image.jpg)
![21. Veryhachium reductum [Pech 1-07: 31,6/94,6] (IC)](https://image.jimcdn.com/app/cms/image/transf/dimension=1920x400:format=jpg/path/s602cffe28acf71e3/image/i40b3f421271fc543/version/1690721357/image.jpg)
Publiziert in:
Zeitschrift Documenta naturae 117 (1998): Seite 35-51, 2 Abbildungen, 2 Tafeln (mit insgesamt 44 Figuren)
- Published online 16-06-2023 by A.M. Heyng -
Geologie, Fazies und Fossilinhalt des Unteren Lias
in der Sandgrube DIETZ bei Pechgraben
(Kulmbach, Oberfranken)
von
Hans-Joachim Gregor & Sepp Hauptmann
Zeitschrift Documenta naturae Bd. 117 / 1998
Zusammenfassung
Es wird ein Profil aus der Sandgrube DIETZ bei Pechgraben nahe Kulmbach beschrieben, das mega- und makrofloristische Funde geliefert hat. Die fluviatile
Fazies war im unteren Lias von dichten Koniferenwäldem begleitet. Ein kleiner Katalog der Megaflora wird mit Zeichnungen des Autors Sepp HAUPTMANN vorgelegt.
Summary
The sandpit DIETZ near Pechgraben (Kulmbach, Northern Bavaria, Germany) yielded abundant remains of a rich rhätoliassic megaflora, mainly with
Podozamites. The limnic-fluviatile conditions of the Sediment, the palaeoambiente and climatological reconstructions are shortly mentioned. Author Sepp HAUPTMANN shows drawings from some
of the most important plants.
Einleitung
Autor GREGOR wurde 1990 von Sepp HAUPTMANN und seiner Frau Traute eingeladen, die Grube DIETZ bei Pechgraben (Oberfranken) zu besuchen. Aus dieser Sandgrube hat das Ehepaar HAUPTMANN einen großen
Teil ihrer berühmten Pflanzensammlung, wohl der größten Lias-sammlung Deutschlands, gewonnen. Ihre Sammlung beinhaltet tausende von Stücken mit einem Reichtum von über 30 Arten allein aus der
Grube DIETZ.
Die Sandgrube DIETZ bei Pechgraben
Die Sandgrube liegt nahe des kleinen Ortes Pechgraben bei Kulmbach (vgl. Abb. 1 des Beitrages FECHNER 1998, in diesem Band) mitten im Wald und lieferte seit Jahren den kleinkörnigen Sand der
mittleren Lagen, wobei die unterlagemden Tone meist als störend empfunden wurden (aus wirtschaftlichen Gründen).
Die Tone bestehen vorwiegend aus Quarz und Kaolinit, wobei nach KESSLER (1973:13) der Illitgehalt im Lias ansteigt. Stratigraphisch dürfte das Sediment in den Unteren Lias gehören, ins Hettangium (vgl. FECHNER 1998, in diesem Band). Aufschlußdaten der Sandgrube DIETZ bei Pechgraben: MTB 1:25 000 No.6135, R44 71 700, H55 41 445, Höhe NN 410-415 m.
Geologie des fränkischen Lias
Zur Geologie sei hier nur kurz erwähnt, daß der früher als "Rhätolias" bezeichnete Fundhorizont fossiler Pflanzen heute als unterster Lias erkannt wurde (JUNG 1960). Es gibt einige Publikationen
zum Thema, speziell von URLICHS 1966,und vor allem von KESSLER 1973, der die verschiedenen fränkischen Horizonte als Rhätolias und Lias bezeichnet hat. Marine Ingressionen liegen im Großbereich
vor, wobei die fluviatilen Schüttungen z.T. weit außerhalb der Küstenbereiche liegen. Der klastische Einflußbereich wird von der Böhmischen Masse her gesehen (vgl. KESSLER,
1973:13).
Die Floren der Grenzschichten wurden bereits von GOTHAN 1914, WEBER 1968 und SCHMEISSNER & HAUPTMANN 1993 gewürdigt.
click to enlarge

Das Profil von Pechgraben
Ein Idealprofil der Grube zeigt Abb. 1, wobei aufgrund der fluviatilen Verhältnisse recht schnelle laterale Fazieswechsel auftreten können, wenn auch die Gesamtkomposition recht einheitlich
bleibt. Basal liegen rötlich-graue Tone vor, dick gebankt, aber gut spaltbar. Diese erbrachten die massenhaften Anreicherungen von Pflanzenfossilien, die in Kap. 3 erwähnt werden. Darüber folgen
diskordant liegend rötlich-gelbliche Sande mit gelegentlichen kohligen Bändchen von 1-2 cm Mächtigkeit und sog. Baumlöchem, d.h. röhrenförmige Überreste von verwesten Baumstämmen. Diese hohlen
Gebilde zeigen nur noch limonitische Krusten, aber keinerlei bestimmbare Innenstrukturen mehr. Sie weisen jedoch in ihrer Vielzahl auf eine dichte Besiedlung der (weiteren) Umgebung hin, auch
wenn man sie als eindeutig allochthon bezeichnen muß.
Im Hangenden folgen fluviatile graue Sande, z.T. kiesig mit erosiver Struktur und lateral wechselnden Mächtigkeiten.
Über zwei kurzfristige Kies-Sand-Lagen mit eingeschalteter Sandlage mit Schräg- und Kreuzschichtung folgen dunkelblaugraue Tone, die z.T. kohlig ausgebildet sind.
Abgeschlossen wird das ganze Profil mit zementierten Bänken von Sandstein (plattig) mit unruhiger Wechsellagerung bis zur Überdeckung mit Waldbodens.
KESSLER hat 1973 (S.51-53) die Profile des fränkischen Rhätolias mit 3 Zyklen in Verbindung gebracht, die partiell als Ingressionen zu deuten sind.
Die Genese des geologischen Systems
Wir haben im Laufe der Zeit eine Abfolge vorliegen, die folgendermaßen zu charakterisieren ist: Eine Zeit der ruhigen, wohl z.T. limnischen Sedimentation (rötliche Tone) in einem dichten
Waldsystem voller Koniferen und Fame wird abrupt durch fluviatile Gegebenheiten gestört und durch Sandlagen überdeckt. Zeitliche Diskordanzen sind sehr wahrscheinlich. Wechselnde
Grundwasserstände, Auskolkungen, Erosionsrinnen, kurzfristige Überflutungen mit Transport von Baumstämmen (später verwest und die "Rinden" als vererzte Schwarten sichtbar) und unruhige
kurzfristige Ruhephasen (Kohlebändchen) kennzeichnen diese wohl kurze fluviatil beeinflußte Zeitspanne.
Nun tritt wieder Ruhe ein, limnische blaugrauerTone werden abgelagert (fossilleer), die wiederum diskordant von Sandsteinen mit schwartenähnlichen Lagen,
bedingt durch wohl episodische Niederschläge, überlagert werden. Eine längere Zeit ziemlich gleichförmigen Transports von Sand schließt die fluviatile Geschichte des Profils ab (vgl. zu allem
KESSLER 1973).
Fossilinhalt des Lias von Pechgraben
Der Fossilinhalt der rötlichgrauen Tone ist als sehr reich zu bezeichnen, wobei vor allem die massenweise vorkommenden Podozamites-Nadeln. (blattähnlich) zu erwähnen sind. Daß an vielen
Nadeln Bißspuren von Insekten zu sehen sind, soll hier akzessorisch erwähnt werden. Solche deutlichen Spuren haben schon GRAUVOGEL-STAMM & KELBER 1996 aus dem Keuper erwähnt.
Auch ein Rest eines Haifisch-Eies wurde gefunden (Palaeoxyris sp.). Aus der benachbarten Grübe KÜFNER liegen ebenfalls Funde vor, die schon früher
publiziert wurden (vgl. KIRCHNER 1992, KIRCHNER & VAN KONIJNENBURG - VAN CITTERT 1994 und SCHMEISSNER & HAUPTMANN 1993).
Palynologische Befunde
Im Beitrag FECHNER in diesem Heft wird die Mikroflora der Grube erschöpfend dargestellt. Als Basis für die Übersicht über die Mikroflora kann die Arbeit von JUNG 1960 gelten, der folgende Formen
festgestellt hat: Sporen vor allem aus den Gruppen Trileites, Horstisporites, Nathorstisporites, Triletes, speziell der Thaumatopteris-Zone (zur
Stratigraphie vgl. ibid.160, 161). Aufgrund des überaus ähnlichen geographischen Verhaltens der diversen liassischen Palynomorphen kam der Autor zum Schluß, daß zu dieser Zeit eine
einheitliche Vegetation im Gebiet des heutigen Europas geherrscht habe (ibid.164). FECHNER hat 1998 (im Beitrag in diesem Band) im Gegensatz dazu andere Formen gefunden, vor allem neben den
Sporen auch die erwarteten Pollenkömer von Koniferen usw. (Cycadopites, Spheripollenites, Taxodiaceaepollenites u.a.) sowie Plankton wie Tasmanites und
Pterosphaeridia, aber auch Innentapeten von Foraminiferen - was wiederum wenigstens einen gewissen marinen Einfluß wahrscheinlich macht.
Die Megaflora
Die Megaflora besteht, abgesehen von Pilzresten auf Podozamites-Blättem (Xylomites zamitae und Xylomites asteriformis), überwiegend aus Teilen von Nadelgewächsen,
Ginkgoverwandten und Farnen, wobei auch ganze Zweige und Wedel auftreten, was wiederum auf parautochtone Verhältnisse in den Tonen hinweist. Auch Schachtelhalmartige, Samenfame und
Cycas-gewächse sind vertreten. Einige schon früher von Autor HAUPTMANN angefertigte Rekonstruktionszeichnungen werden hier als Überblick zur Flora gebracht. Im Einzelnen sind folgende
höhere Pflanzenformen zu erwähnen (Liste erstmals von Frau Traute HAUPTMANN zur Verfügung gestellt):
Pteridophytina
Equisetopsida
Neocalamites lehmannianus - Abb.2
Equisetites muensteri - Abb. 3
Pteridopsida
Selenocarpus muensterianus - Abb.5
Phlebopteris muensteri - Abb.4
Phlebopteris angustiloba
Phialopteris tenera
Thaumatopteris brauniana
Thaumatopteris schenki
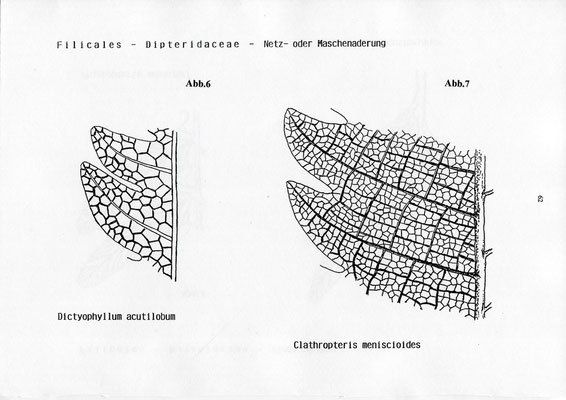
Clathropteris meniscioides - Abb.7
Dictyophyllum nilssoni (hon acutilobum, schriftl. Mitt. v. KONIJNENBURG) - Abb.6
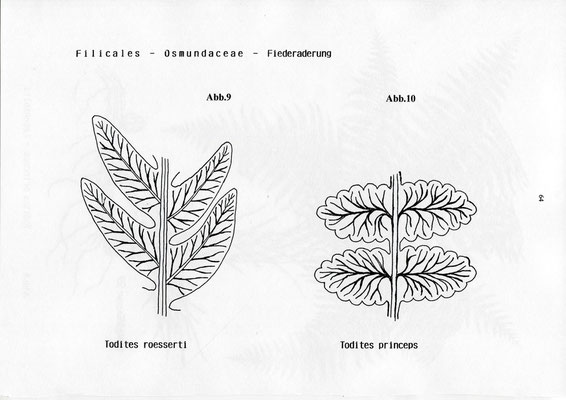
Todites princeps - Abb. 8,10
Todites roessertii - Abb.9
Coniferophytina
Pinopsida
Hirmerella muensteri - Abb. 12
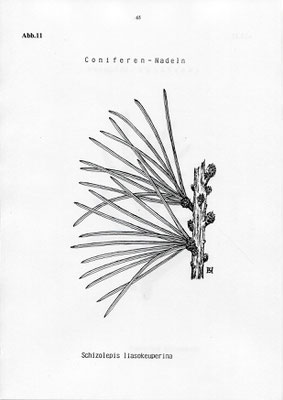
Schizolepis liasokeuperina - Abb. 11
Podozamites distans - Abb. 13
Swedenborgia benkerti
Swedenborgia cryptomerides
Swedenborgia major
Ginkgoopsida
Baiera muensteriana
Karkenia hauptmannii (mit Zapfen)
Schmeissneria microstachys (Einzelblätter)
Stachyopitys presslii
Cycadophytina
Pteridospermae
Umkomasia franconica
Thirmfeldia rhomboidalis
Cycadopsida
Nilssonia acuminata
Nilssonia polymorpha
Gnetopsida
Desmiophyllum gothani
Chlamydolepis lautneri
Bemettia inopinata
Piroconites keusperti
indet.
Reste von Rhizomopteris und Carpolithes sp.
Einige weitere Reste von Pechgraben, aber nicht speziell aus der Grube DIETZ, sondern auch besonders aus der Grube KÜFNER sollen hier vorläufig erwähnt werden. Eine gesamte Bearbeitung des
Florenkomplexes steht noch aus:
? Palissia sphenolepis (Coll. SCHMEISSNER)
Campylophyllum hörmannii (Coll. SCHMEISSNER)
Marattiopsis intermedia (Coll. v.KONUNENBURG)
Pseudoctenisprossii (Coll. SCHMEISSNER)
Cycadites sp. (Coll. v.KONUNENBURG)
Cycadospadix sp. (Coll. v.KONIJNENBURG)
Nilssoniopteris cf. vittata (Coll. v.KONIJNENBURG)
Otozamites brevifolius (Coll. v.KONIJNENBURG)
Weltrichia mirabilis (Coll. v.KONIJNENBURG)
Eine weitere reiche Flora aus der Nähe von Coburg, aus dem dortigen Rhät, erwähnen KELBER &, VAN KONIJNENBURG - VAN CITTERT 1997. Ein vorläufiger Vergleich des basalen Haupttones von
Heilgersdorf mit der Tonlinse von Pechgraben liefert folgendes Ergebnis (Tab. 1):
click to enlarge
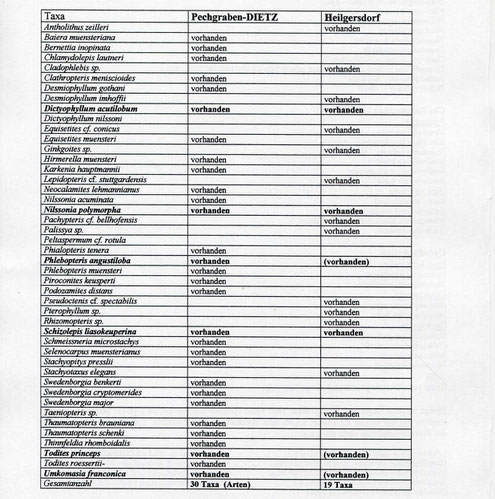
Tabelle 1:
Vergleich der Floren von Pechgraben (Lias) und Heilgersdorf (Rhät) - in Klammer die nächstverwandte Art).
Man sieht sehr deutlich, daß schon in der kleinen Zeitspanne Rhät zu Lias ein gewaltiger Wechsel in der Flora auftritt - auch wenn die Fazies etwa gleichgeblieben
ist. Gemeinsam sind den beiden Fundstellen ja nur 6 Taxa, wobei nur 3 den Artenvergleich gestatten (Fettdruck in Tabelle 1). Weitere Studien in dieser Richtung könnten interessante Ergebnisse zu
ökologisch-stratigraphischen Fragestellungen ergeben, denn hier ist schon eindeutig ein stratigraphischer Unterschied zu sehen.
click to enlarge & read figure captions





Paläoambiente
Wenn wir versuchen, eine Rekonstruktion des ökologischen Systems von Pechgraben zu machen, kommen wir bei den linsenförmigen Tonkörpem auf Ablagerungs-Bedingungen, wie sie in seichten
Rinnensystemen oder Altwasserbereichen großer Ströme vorliegen. Wir müssen zwar die Sandsteinbereiche hiervon insofern abtrennen, als diese den Hauptbereich der Flüsse darstellen, können aber das
Gesamtsystem mit diversen Meeresvorstößen und Rückzügen im Übergang Trias-Jura koppeln (KESSLER 1973:51).
Schnelle Sedimentaton erscheint typisch für die rötlich-grauen Tone, da die Pflanzen hervorragend erhalten sind, keinerlei Einrollungen verkommen oder Verwesungsspuren oder Pilze sichtbar sind. Im Gegensatz dazu müssen bei den Baumstämmen im Sandstein lange Zeit gut durchlüftete Verhältnisse vorgelegen haben, da sie praktisch vollkommen verwest sind und nur noch die röhrenförmigen Ummantelungen der Stämme vorliegen.
Klimarekonstruktion
Wie schon KESSLER (1973:14) erwähnte, dürfte das Klima zur Zeit der reichen Pechgrabenflora als humid und subtropisch, evtl, tropisch anzusehen sein. Hohe Niederschläge und ebensolche
Temperaturen bedingen die typische Tonverwitterung, die reiche Flora und die sedimentologischen Gegebenheiten wie point bar- oder Altwasserlagen. Besondere Merkmale wie Spalt-öffnungen usw.
müßten erst noch weiter untersucht werden, um nähere Aussagen zu gestatten. Daß sich dabei aber auch manchmal fehlinterpretierte Bedingungen ergeben, zeigen diverse spezielle Arbeiten, die mit
dem Spaltöffnungsmodus fossiler Pflanzen, ein aus anderen Gründen gefordertes arides Klima verneinen.
FECHNER hat wiederum (1998, in diesem Band) die Daten für die genannten Klimadaten voll bestätigt mit „humid und warm-gemäßigt (subtropisch)“ und überträgt sie somit in das Hettangium, also den
Unteren Lias.
Literatur
FECHNER, G.G. (1998): Eine Mikroflora aus Pechgraben und ihre stratigraphische Stellung im Lias von Franken (S-Deutsch-land). - Documenta naturae, 117: 35-51, 2 Abb., 2 Taf., München.
GOTHAN, W. (1914): Die unterliassische (rhätische) Flora der Umgegend von Nürnberg. - Abh. NaturhistGes. Nürnberg. - 19: 89 - 186, Nürnberg.
GRAUVOGEL-STAMM, L. & KELBER, K.-P. (1996): Plant-insect interactions and coevolution during the Triassic in Western Europe.- Paleontologica Lombardia, N. S. 5: 5-23; Milano.
JUNG, W. (1960): Die dispersen Megasporen der fränkischen Lias-Grenzschichten. - Palaeontographica£, 107:127-170, Taf. 36-39, Stuttgart.
KELBER, K.-P, & VAN KONUNENBURG-VAN CITTERT, J.H.A. (1997): A new Rhaetian flora from the neighbourhood of Coburg, (Germany) - preliminary results.- Proceedings 4th. Europ. Palaeobot.
Palynol. Conf. Heerlen 1994 (EPPC); Meded. Ned. Inst. Toegepaste Geowetenschappen TNO, 58:105-113; Haarlem.
KELBER, K.-P. (1998): Phytostratigraphische Aspekte der Makrofloren des süddeutschen Keupers- Documenta naturae, 117: 1-11,2 Abb., 2 Taf, München.
KESSLER G. (1973): Sedimentgeologische Untersuchungen im oberfränkischen Rhätolias.- Erlanger Geol. Abh., 93:1-60,20 Abb., 4 Taf., 6 Diagr., Erlangen.
KIRCHNER, M. & VAN KONIJNENBURG - VAN CITTERT, J. H A. (1994): Schmeissneria microstachys (PRESL 1833) KIRCHNER & VAN KONIJNENBURG - VAN CITTERT comb. nov. and Karkenia
hauptmanni KIRCHNER & VAN KONIJNENBURG - VAN CITTERT sp. nov., plants with ginkgolean affinities from the Liassic of Germany. - Rev. Palaeobot. Palynol., 83, 1 - 3: 199 - 215;
Amsterdam.
KIRCHNER, M. (1992): Untersuchungen an einigen Gymnospermen der fränkischen Rhät - Lias -Grenzschichten. - Palaeontographica, B, 224,1 - 3: 17 - 61; Stuttgart.
SCHMEISSNER, S. & HAUPTMANN, S. (1993): Fossile Pflanzen aus den Rhät - Lias - Übergangsschichten des Kulmbach - Bayreuther Raumes. „Glossophyllum“ - ein 200 Millionen Jahre alter
Baum. - Naturwiss. Ges. Bayreuth, Ber. XXH: 51-66; Bayreuth.
SCHMEISSNER, ST. & HAUPTMANN, S. (1998): Ein Blattschopf von Nilsonia acuminata (PRESL) GOEPPERT aus dem Unteren Lias Oberfrankens.- Documenta naturae, 117: 1-11, 2 Abb., 2 Taf.,
München.
URLICHS, M. (1966): Zur Fossilführung und Genese des Feuerlettens, der Rät - Lias - Grenzschichten und des unteren Lias bei Nürnberg. - Erlanger geol. Abh.
64: 1 - 42; Erlangen.
VAN KONIJNENBURG - VAN CITTERT, J. H. A., SCHMEISSNER; ST. & HAUPTMANN; S. und T. (1998): Neue Ergenbisse zu Ctenozamites wolfiana (Pteridospermae) und Pseudoctenis
prossii nov.spec. (Cycadophyta) aus dem Unteren Lias (Jura,Bayern). - Documenta naturae, 117: 13-33,6 Taf., München.
WEBER, R. (1968): Die fossile Flora der Rhät - Lias - Übergangsschichten von Bayreuth (Oberfranken) unter besonderer Berücksichtigung der Coenologie. - Erlanger geol. Abh. 72: 1-73;
Erlangen.
- Published online 20-12-2023 by A.M. Heyng -
click to enlarge

Tafel 1
Fig. 1: Ansicht der Grube DIETZ vom Waldweg aus.
Fig. 2: Gesamtprofil in der Grube, die fluviatilen Rinnen und Strukturen zeigend.
Fig. 3: basaler Teil der Grube DIETZ mit rötlich-grauen Tonen und hangenden fluviatilen Sanden.

Tafel 2
Fig. 1: hangende fluviatile Sande und Kiese mit graublauen Tonlagen unter gelbgrauen zementierten Sandsteinen.
Fig. 2: Sepp Hauptmann 1990, beim Freilegen von großen Sedimentplatten mit Podozamiten.
Fig. 3: basale rötlichgraue Tone mit Podozamites-Fossilien.
Fig. 4: rötliche Tone mit hangenden streifigen Sanden.
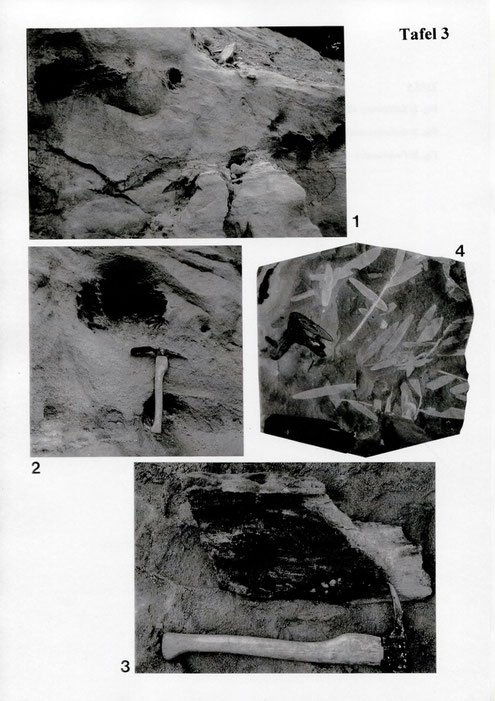
Tafel 3
Fig. 1: Fluviatile Sande mit Baumstamm-Löchern.
Fig. 2: Einzelne "Stammlöcher" im grauen Sand.
Fig. 3: Vererzte Reste von großen Baumstämmen.
Fig. 4: Sedimentplatte (roter Ton) mit Anhäufung von Podozamites distans.

Tafel 4
Fig. 1: Kohlentone als Rinnenfullung im rötlichgrauen Ton der Basis.
Fig. 2: Sedimentstruktur im Sand.
Fig. 3: Podozamites-Nadel mit Insektenbiß (vgl. auch GRAUVOGEL-STAMM & KELBER 1996).
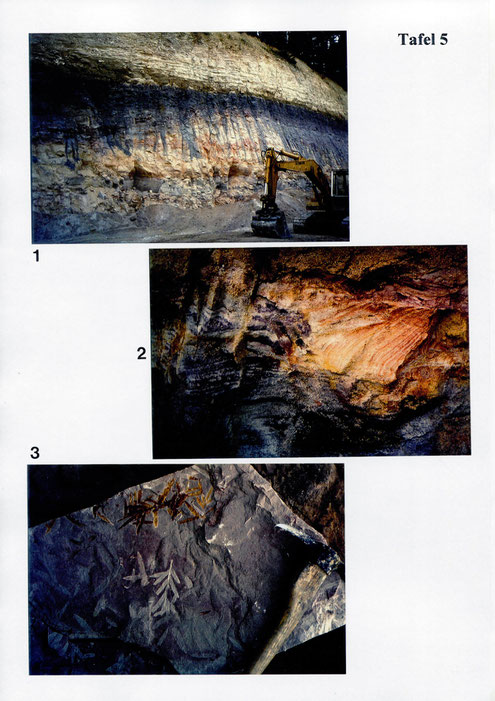
Tafel 5
Fig. 1 : Aufschlußwand mit basalem rötlichen Ton, farbiger Serie in graublauen Tonen im Hangenden,
überdeckt mit beigegrauen Tonen.
Fig. 2: fluviatile Sedimentstruktur mit farbigen Lösungs-erscheinungen.
Fig. 3: rötliche Tonplatte mit Podozamites-Zweigen und - Nadeln; z.T. bräunlich-gelb durch Eisenlösungen verfärbt.
- Weiterführende Informationen zum Thema Pechgraben -
Phytostratigraphische Aspekte der Makrofloren
des süddeutschen Keupers
von
Klaus-Peter Kelber
Zeitschrift Documenta naturae Bd. 117 / 1998
Abstract
Four prominent fossil macroplant zones can be distinguished in the Keuper (Upper Triassic) of Southern Germany: Lower Keuper Flora ("Lettenkeuper", Ladinian), Schilfsandstein Flora (Carnian), Coburger Sandstein Flora (Upper Carnian?/Norian?) and the Rhaetian Flora. Only a few fossil plant remains from the Gipskeuper (Carnian) and the Stubensandstein/Burgsandstein (Norian) support the present state of palaeobotanical knowledge.
Patterns in vascular land plant diversification of the Keuper are characterized by distinctive taxa that first appeared in the Triassic (modern mesophytic floral elements) and some survivors from the Palaeozoic (palaeophytic floral elements).
Lower Keuper and Schilfsandstein taphofloras from many localities in Southern Germany are mainly represented by horsetails of which Equisetites arenaceus forms the most dominant floral component. Palaeophytic floral elements are the equisetalean genera Schizoneura and Phyllotheca, the marattialean fern Danaeopsis, "Voltzia"-type conifers, the cycads and some Pteridospermopsida, particularly Lepidopteris and its fructifications. The salient evolutionary feature is the appearance of Bennettitales as testified by Pterophyllum from Lower Keuper and later floras, and culminating in the Schilfsandstein Flora. Modern leptosporangiate ferns of the genera Dictiophyllum and Clathropteris (Dipteridaceae) occur in the Schilfsandstein Flora for the first time. New finds indicate that conifers became highly diverse in the Coburger Sandstein plant beds. The sparsely distributed Rhaetian floras display a major shift to modern mesophytic floral composition. This flora includes Phlebopteris, Pachypteris ["Thinnfeldia"], Umkomasia, Nilsonia, Schizolepis, Stachyotaxus, and Palissya. "Voltia"-type conifer elements and the peltasperm Lepidopteris disappear at the end of the Triassic.
New finds, especially from the Lower Keuper Flora (Schizoneura, Phyllotheca, Linguifolium, reticulate venation pattern, ?"Glossopteris", ?"Sagenopteris") suggest more uniform floral components between Euramerica and Gondwana than previously assumed. Phytostratigraphical work on macroplants in the Keuper Flora of Southern Germany is currently handicapped by insufficient taxonomic research as well as sampling deficiency for many Keuper sequences.
Concerning the evaluation of the end-Triassic floral mass extinction event and the subsequent vegetational recovery a “golden spike" situation is located adjacent to Coburg and Bayreuth in Northern Bavaria. Plant bearing localities of the former indistinguishable "Rhät-Lias-Grenzschichten" now have been properly defined as the Rhaetian and the successive Liassic plant beds on sedimentological and palynological grounds. Obviously there is no disruption or decline in species diversity neither in Rhaetian nor in Liassic taphofloras in this area.
Einleitung
Bei annähernd konstanter Palaogeographie des mitteleuropäischen Triasbeckens konnte sich im Keuper (Oberes Ladin bis Rhät) über einen Zeitraum von annahernd 24 Millionen Jahren mit wenigen Unterbrechungen eine terrestrische Flora fortentwickeln. Vier markante Florenkomplexe - im Unterkeuper, Schilfsandstein, Coburger Sandstein (Kieselsandstein) und Rhät - geben gegenwärtig Auskunft über diese Entwicklung. Nur sehr wenige sporadische Pflanzenfunde aus den übrigen Schichtgliedern des Keupers, vorwiegend aus dem Gipskeuper und dem Burgsandstein (Stubensandstein), können bislang dieses paläöbotanische Wissensbild der süddeutschen Keuperfloren ergänzen (Abb. 1).
Abbildung 1. Stratigraphische Übersicht des süddeutschen Keupers. Nach Geyer & Gwinner (1991), verändert.
Die reichhaltigsten Pflanzenfossil-Aufsammlungen des süddeutschen Keupers liegen aus dem Unteren Keuper (Hauptsandstein- oder Werksandstein-Bereich) und aus dem Schilfsandstein vor (Abb. 2). Bemerkenswert ist nach einer weltweiten Phase aussetzender oder geringerer Kohlebildung zwischen Unter- und Mittel-Trias (Retallack et al. 1996) das reichliche Vorkommen von lokalen Kohleflözen im Unteren Keuper. Deren paläobotanische Erforschung steht allerdings noch aus, ebenso die Beschreibung der jüngsten Bernstein-Funde aus dem Schilfsandstein (Kelber & Hansch 1995: 89). Hervorzuheben sind in diesem Zusammenhang auch die ehemals durch Pyrit permineralisierten Pflanzenteile aus dem Unteren Keuper, die ein bis heute noch kaum ausgewertetes Reservoir strukturbietender Information repräsentieren (Brunner & Kelber 1988). Spärliche Funde der Interaktion Tier/Pflanze (Besiedelungsspuren von Spirorbis sp., Eigelege und FraBspuren von Insekten) konzentrieren sich auf das Ökosystem des Unteren Keupers (Grauvogel-Stamm & Kelber 1996). Sie vermitteln einen willkommenen Einblick in das komplexe Beziehungsgeflecht der damaligen Lebewelt und deren Beziehungen zur unbelebten Natur.
Bei weltweiter Betrachtung blieben in mesozoischen Floren vor allem die Blattreste von Cycadeen und Bennettiteen überliefert (Taylor & Taylor1993). Diese treten in der Unterkeuper- und Schilfsandsteinflora quantitativ jedoch nur untergeordnet oder nur in Form disperser Kutikeln in Erscheinung. Sie sollen aus dem fränkischen Schilfsandstein teilweise in einer besonderen Erhaltungsform zusammen mit Resten fossiler Flechten überliefert worden sein (Ziegler 1997). Von zahlreichen Unterkeuper- und Schilfsandstein-Fundorten Süddeutschlands wurden jedoch ausschlieBlich nur monotone Taphofloren geborgen, die durch Equiseten, insbesondere Equisetites arenaceus (Kelber & van Konijnenburg-van Cittert 1998), E. conicus und Neocalamites merianii dominiert sind. Das gehäufte Vorkommen solcher krautigen Equisetales ist auch in globaler Hinsicht in den fossilen Floren nur sehr selten verwirklicht und harrt noch einer phytotaphonomischen Erklärung.
Der palaobotanische Wissensstand über viele Florenelemente des süddeutschen Unterkeupers und Schilfsandsteins resultiert aus der Bearbeitung der Keuperflora von Lunz in Österreich und von Neue Welt bei Basel. Diese Florenkomplexe werden als zeitgleich mit dem Schilfsandstein korreliert.
Abbildung 2.
Position der Florenkomplexe in einer vereinfachten maßstabgetreuen stratigraphischen Übersicht des Keupers (nach Menning, 1995), sowie geographische Lage einiger Fundorte von Keuperpflanzen in Süddeutschland (nach Kelber & Hansch 1995, verändert).
Coburger Sandstein: Die ergiebigsten Aufsammlungen aus diesem stratigraphischen Horizont beschränken sich bislang auf die Steinbrüche des "Weißen Mainsandsteins" im Bereich von Zeil und Ebelsbach (Abb. 2). Es herrscht der Eindruck einer durch Koniferen dominierten Taphoflora (sterile Zweige, Steinkernerhaltung von Koniferenholz). Durch Prospektion und Ausgrabung einer neuen Fossil-Lagerstätte von Triops cancriformis (Branchiopoda, Notostraca, s. Kelber, im Druck) konnten im Nebeneffekt auch fossile Pflanzenreste geborgen werden, die das bisher bekannte Florenspektrum erweitern. Neufunde sind Equisetites arenaceus, Pterophyllum brevipenne, cycadeen- oder bennettiteen-artige Fruktifikationen, Elatocladus sp. (Taf. 2, Fig. 2), cf. Eoginkgoites sp. (Taf. 2, Fig. 6), cf. Czekanowskia sp. (Taf. 2, Fig. 5), Desmiophyllum imhoffi, "Albertia" sp. (Taf. 2, Fig. 10). Einen besonderen Informationsspeicher mikroskopischer Baupläne stellen die zahlreichen Holzreste dar, die in Fussit-Erhaltung vorliegen. Darüber hinaus bezeugen diese Reste Wildfeuer in der Paläovegetation des Keupers.
Rhät: Die wenigen echten Rhätfloren des süddeutschen Keupers beinhalten die bisher einzige Taphoflora mit gröBerer Diversität und ohne ausgesprochene Dominanz eines oder mehrerer Florenelemente. Als bekanntester Fundort gilt, neben den baden-württembergischen Vorkommen und dem von Burgpreppach (Taf. 3, Fig. 6), die Lokalitat Kipfendorf bei Coburg (Taf. 3, Fig. 3, 4), von der auch eine revidierte Florenliste der Makro-Pflanzenreste vorliegt (Jung in Achilles 1981). Der neu entdeckte Fundpunkt Heilgersdorf bei Sesslach (Kelber & van Konijnenburg-van Cittert 1997) ergänzt das Spektrum durch folgende Elemente: Equisetites, Phlebopteris, Pachypteris ["Thinnfeldia"], Umkomasia (Taf. 3, Fig. 5), Peltaspermum (Taf. 3, Fig. 7), Antholithus, Pseudoctenis, Nilssoniopteris, Nilsonia (Taf. 3, Fig. 2), Ginkgoites, Stachyotaxis (Taf. 3, Fig. 1), Geinitzia vel Elatides, Schizolepis, Palissya.
Insgesamt vermitteln die süddeutschen Rhätfloren den Eindruck einer noch etwas verarmten Liasflora, bedingt nicht zuletzt durch den mangelhaften Stand der Aufsammlungen. Im Kontrast zu den älteren Unterkeuper-, Schilfsandstein- und Coburger Sandstein-Floren sind die wichtigen Gruppen der Matoniaceen, Dipteridaceen und Nilsoniaceen in voller Höhe entwickelt. Dieses gleichförmige Florenbild, mit noch etwas stärkerer Beteiligung der Bennettiteen und Ginkgophyten, bleibt dann weltweit kennzeichnend für alle Jura-Floren.
... weiterlesen im pdf - freier Download, open
access - siehe unten >>>
Tafel 1
Maßstab, soweit nicht anders bezeichnet, jeweils 1 cm. (OCH-, ROT-, SCHL- = Coll. des Verfassers).
Fig. 1. Danaeopsis arenacea, Fiederfragment. Unterer Keuper, Buchbrunn bei Kitzingen. Pal. Mus. Humboldt Univ. Berlin, (o. Nr.). Abbildungsoriginal
zu Schoenlein 1865, Taf. 7, Fig. 3.
Fig. 2. Swedenborgia sp., Zapfenschuppe. Unterer Keuper, Schleerieth, SCHL-807. Vergrößerung: x 5,6.
Fig. 3. Equisetites arenaceus, Sproßgipfel, 12,5 cm Durchmesser. Unterer Keuper, Wolfgangsberg bei Ochsenfurt, OCH-007a.
Fig. 4. Chiropteris digitata, Wedelfragment mit Netznervatur. Unterer Keuper, Estenfeld bei Würzburg. Pal. Mus. Humboldt Univ. Berlin, Nr. 1989/491/3745. Abbildungsoriginal zu Schoenlein 1865, Taf. 11, Fig. 1a.
Fig. 5. Fruchtblatt einer noch nicht näher bestimmten Cycadee (distaler Bereich wie bei "Dioonitocarpidium"). Unterer Keuper, Schleerieth,
SCHL-038a.
Fig. 6. Schizoneura paradoxa, beblätterte Achse. Schilfsandstein, Rothof bei Oberlauringen, ROT-028a.
Fig. 7. Annalepis zeilleri, Sporophyll. Unterer Keuper, Lettenkohlendolomit (Lingulabänke), Rottmünster (gemeint ist wahrscheinlich Rottenmünster,
ein Stadtteil von Rottweil). Pal. Mus. Univ. Tübingen. Erste photographische Darstellung des Abbildungsoriginals zu Quenstedt 1885, Taf. 95, Fig. 19. Ursprünglich von Quenstedt als Nagelblatt
"Onychophyllum" bezeichnet, geriet dessen Abbildung und Beschreibung jedoch vollkommen in Vergessenheit. Erst 1910 wurden gleichartige Fossilien als Annalepis
zeilleri erneut beschrieben.
Fig. 8. Linguifolium sp., Fiederfragment. Unterer Keuper, Weiler zum Stein. Staatl. Mus. Naturkde. Stuttgart, P. 1740/1.
Fig. 9. Phyllotheca sp., Blattscheidenfragment. Unterer Keuper, Schleerieth, SCHL-029.
Fig. 10. "Sagenopteris" sp., Fiederfragment mit Netzaderung. Unterer Keuper, Weiler zum Stein. Staatl. Mus.
Naturkde. Stuttgart, P. 1739/1.
click to enlarge & read figure captions








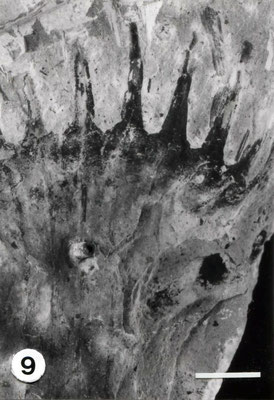

Tafel 2
Maßstab jeweils 1 cm. (ELT-, PASS-, Z- = Coll. des Verfassers)
Fig. 1. Pterophyllum jaegeri, geknickter Wedel mit Wedelspitze. Schilfsandstein, Zeil am Main, Z-012.
Fig. 2. Elatocladus sp., beblätterter Zweig. Coburger Sandstein, Steinbruch an der Passmühle, Ebelsbachtal, PASS-056a.
Fig. 3. Voltzia coburgensis, gegabelter Zweig. Coburger Sandstein, Eltmann, ELT-013.
Fig. 4. Dictyophyllum exile, Wedel. Schilfsandstein, Stuttgart. Staatl. Mus. Naturkde. Stuttgart, P. 778/3.
Fig. 5. cf. Czekanowskia sp., Blattsegment. Coburger Sandstein, Steinbruch an der Passmühle, Ebelsbachtal, PASS-060.
Fig. 6. cf. Eoginkgoites sp., Blattsegment. Coburger Sandstein, Eltmann, ELT-068.
Fig. 7. Widdringtonites keuperianus ("Brachyphyllum"-Habitus), mit Abdrücken der in Reihen angeordneten Spaltöffnungen. Coburger Sandstein, Steinbruch an der Passmühle, Ebelsbachtal, PASS-055b.
Fig. 8. Voltzia coburgensis, Zapfenschuppe. Coburger Sandstein, Ziegelanger bei Zeil am Main. Coll. Kehl, Geol. Inst. Univ. Erlangen. Abbildungsoriginal zu Rutte & Wilczewski 1983, Taf. 2, Fig. 6.
Fig. 9. "Glyptolepis" platisperma, Zapfenschuppe. Coburger Sandstein, Steinbruch an der Passmühle, Ebelsbachtal, PASS-057.
Fig. 10. "Albertia" sp., beblätterter Zweig. Coburger Sandstein, Steinbruch an der Passmühle, Ebelsbachtal, PASS-058a.
click to enlarge & read figure captions





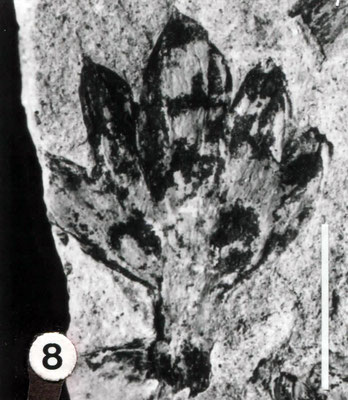

Tafel 3
Maßstab jeweils 1 cm. (HEI- = Coll. des Verfassers))
Fig. 1. Stachyotaxis elegans, beblätterter Zweig. Rhät, Heilgersdorf bei Sesslach, HEI-070a.
Fig. 2. Nilsonia sp., Blattfragment. Rhät, Heilgersdorf bei Sesslach, HEI-O25.
Fig. 3. Phlebopteris muensteri, Wedelbasis. Rhät, Kipfendorf bei Coburg. Pal. Mus. Humboldt Univ. Berlin, Nr. 381. Abbildungsoriginal zu Mägdefrau 1968, Abb. 247a.
Fig. 4. Lepidopteris ottonis, Wedel, mit Zwischenfiedern an der Rhachis (Pfeile). Rhät, Kipfendorf bei Coburg. Pal. Mus. Humboldt Univ. Berlin, Nr. J.1729. Nach Angaben auf dem
Fundzettel aufgesammelt durch Loretz, 1893.
Fig. 5. Umkomasia sp., Fragment einer Blüte. Rhät, Heilgersdorf bei Sesslach, HEI-211a.
Fig. 6. Clathropteris meniscioides, Blattfragment. Rhät, Burgpreppach. Coll. Kehl, Geol. Inst. Univ. Erlangen. Abbildungsoriginal zu Rutte & Wilczewski 1983, Taf. 2, Fig. 10.
Fig. 7. Peltaspermum sp., weibliche Blüte von Lepidopteris ottonis. Rhät, Heilgersdorf bei Sesslach, HEI-232.
Fig. 8. Dictyophyllum acutilobum, Fieder. Rhät, Heilgersdorf bei Sesslach, HEI-169.
Fig. 9. Lepidopteris ottonis, Fiederchen in Substanzerhaltung. Rhät, Heilgersdorf bei Sesslach, HEI-086.
click to enlarge & read figure captions









Publiziert in:
Zeitschrift Documenta naturae 117 (1998): Seite 89-115, 3 Abbildungen, 3 Tafeln

- Published online 15-06-2023 by A.M. Heyng -
Ptilophyllum maculatum GIVULESCU 1992 or Ptilophyllum
aninaenis CZIER 1995 - a problem of priority
by
Razvan Givulescu
Zeitschrift Documenta naturae Bd. 117 / 1998
Zusammenfassung
Der Verfasser behandelt ein Prioritätsproblem, und zwar handelt es sich einerseits um Ptilophyllum maculatum GIVULESCU 1992, andererseits um Ptilophyllum aninaensis CZIER 1995, beide vom selben Fundort - Anina / ante Steierdorf / - Banat, Rumänien. Anhand eines Vergleichs der beiden Originalbeschreibungen und von Photos demonstriert der Verfasser, daß beide "Arten" dieselbe kutikulare Struktur besitzen, daß es also zwischen ihnen keinen Unterschied gibt. So ist der gültige Name des betreffenden Fossils Ptilophyllum maculatum GIVULESCU 1992.
Abstract
A problem of nomenclature priority between Ptilophyllum maculatum GIVULESCU 1992 and Ptilophyllum aninaensis CZIER 1995, both from Anina (Romania) is presented. The author shows that there are no differences between the named "species" in the anatomical structure of the two cuticles, so the material shall be called Ptilophyllum maculatum GIVULESCU 1992.
... weiterlesen im pdf - freier Download, open access >>>
Publiziert in:
Zeitschrift Documenta naturae 117 (1998): Seite 81-87, 2 Plates

- Published online 15-06-2023 by A.M. Heyng -