
Projekt Plattenkalk
News & Aktuelles
_____________________________________________________________________________________________
Historische Handstücke zur Plattenkalk-Fazies
Published online 13-01-2022 by A. M. Heyng
















"2068. Kalkstein. halb gelb, halb grauer Schiefer von Bietenfeld (= Pietenfeld) b. Eichst. 1. 3. a" (Originalbeschreibung)
Historisches Stück aus der ehemaligen Lehrsammlung des Königlichen Schullehrer-Seminars in Eichstätt; wiss. Slg. amh-Geo.
<< please click on pictures to enlarge




























"2095. Kalkschiefer mit Ocker gefärbt. Eichstaett. A I 1, 3 a bb" (Originalbeschreibung)
Historisches Stück aus der ehemaligen Lehrsammlung des Königlichen Schullehrer-Seminars in Eichstätt; wiss. Slg. amh-Geo.
<< please click on pictures to enlarge
____________________________________________________________
Aus dem Archiv der Documenta naturae Sonderbände (Band 54, 2008)
Ammoniten-Drehmarken aus den Solnhofener Plattenkalken – Befund und Deutung
A.E. Richter*
*Andreas E. Richter, Ifenstr. 8 1/2, 86163 Augsburg, Germany
(Auszug aus der Publikation)
3 Zusammenfassend folgende Aussagen:
1) Herkunft: Die Platte stammt aus dem Plattenkalkgebiet der Südlichen Frankenalb/Bayern. Die Herkunft aus Painten ist wenig wahrscheinlich; die Herkunft aus Zandt müsste überprüft werden.
2) Stratigraphische Stellung: Weißjura epsilon oder zeta (Oberjura; oberstes Kimmeridge bis Untertithon).
3) Verursacher: Die Marken wurden verursacht durch ein komplettes Ammonitengehäuse mit weitestgehend oder komplett erhaltenenen Apophysen.
4) Der Ammonit gehört zu den Perisphinctaceae. Die Gattungs-Zugehörigkeit kann nicht ermittelt werden. Es kann sich um einen Vertreter der Gattungen/Untergattungen Virgataxioceras
(Ataxioceratidae, Ataxioceratinae), Lithacoceras (Subplanites; Ataxioceratidae, Lithacoceratine), Usseliceras (Subplanitoides; Ataxioceratidae,
Lithacoceratine), Parapallasiceras (Ataxioceratidae, Lithacoceratine), Lemencia (Ataxioceratidae, Lithacoceratine) gehandelt haben. Mikrokonche dieser Gattungen/Untergattungen
wurden in den Schichten der
Solnhofen-Formation nachgewiesen.
5) Die Marken entstanden in einem zur Bildungszeit praktisch unbewegten Lagunenbecken mit allergeringsten Strömungseinwirkungen. Wie der Ammonit in das Lagunenbecken kam, ist nicht zu klären. Ein
lebenslanger Aufenthalt hier ist nicht möglich gewesen. Eine Einspülung über die Riffbarren hätte eine Bewahrung der immer hauchdünnen und höchst empfindlichen Apophysen normalerweise unmöglich
gemacht. Also ist anzunehmen, dass das Tier durch ein Barrentor aus einem verträglichen Habitat einwanderte, den Rückweg nicht mehr fand und aufgrund der ungünstigen Umweltbedingungen in der
Lagune starb.
Um auf einer plastischen Tonplatte Versuche zu den Mündungsabdrücken machen zu können, wurden zwei Ammoniten entsprechend vorbereitet. Bei zwei perisphinctiden Ammonitengehäusen aus dem Oxford
(unterer Oberjura) Madagaskars wurde die Mündung auf einen Tiefe von rund 1,5 Zentimetern als Röhre freipräpariert. Dabei handelt es sich um das apophysentragende Gehäuse eines
Divisosphinctes besiairei COLLIGNON (Durchmesser zehn Zentimeter) mit linksseitig röhrenwurmbewachsener Apophyse und einer schwächer und sehr fein
ausgebildeten rechten Apophyse. Beim zweiten etwas kleineren Gehäuse (Durchmesser 6,8 Zentimeter) handelt es sich um die Innenwindungen eines Exemplars vermutlich der gleichen Art.
4 Die Abdrücke
Die auf diesem Weg mit den madagassischen Perispinctiden auf der Tonplatte erzeugten Marken gleichen den Marken auf der Fossilplatte vollkommen, abgesehen von Feinheiten wie Eindruckstiefe und
Drehwinkel. Ich versuche eine knappe Analyse dieses mir bisher unbekannten Markentyps: Das Ammonitengehäuse driftete knapp über dem Meeresboden, in senkrechter Lage gehalten durch Gasreste im
Phragmokon oder Verwesungsgase in der Wohnkammer. Die Lage im Wasser war so, dass sich die Mündung knapp über dem Meeresboden befand. Grundsätzlich war eine Stabilität in der Medianrichtung nicht
gegeben, da der Schwerpunkt etwa gehäusemittig lag, so dass ein Überkippen in Medianrichtung leicht möglich war und öfters stattfand. Beim Verdrehen um die Vertikalachse verursachte das Gehäuse
jeweils nahezu
gerade bis stark gebogene Rillen entsprechend der Venterberippung, woduch Drehmarken entstanden. Aufgrund der hier erkennbaren Berippung kann der Ammonit in die Gruppe der Perisphinctiden
eingegliedert werden. Das Gehäuse wurde nicht nur linear versetzt, sondern es pendelte auch in Richtung Medianebene. Dabei erfolgten Überkippungen, die am Abdruck der Mündung erkennbar sind. In
der Regel pendelte das Gehäuse nach dem Mündungsaufsatz wieder zurück, da ein die vollkommene Verkippung anzeigender Ventereindruck fehlt.
Neben den Rippenmarken erkennt man vielfach Apophyseneindrücke, teilsweise nur einseitig (Schräglage?). Die kurzen leicht keilartigen Eindrücke neben den Marken weisen meist in die lineare
Hauptrichtung der Marken und stammen von Apophysen des Gehäuses. Diese Eindrücke wurden beim Pendeln durch jeweils kurzes Auftippen des ansonsten knapp über dem Substrat schwebenden
Mündungsbereichs
verursacht. Bei geringstreckigem Versatz entstanden auf diese Art Repetitionsmarken. Ob das Gehäuse nun Mündung "vorn" oder Mündung "hinten" durchs Wasser driftete, ist ohne entsprechende
Versuche nicht zu klären. Apophysenerhaltung bei Plattenkalk-Perisphinctiden ist grundsätzlich sehr selten. In den Mörnsheimer Bankkalken kommen Ammoniten mit erhaltener Mündung und Apophysen
relativ häufig vor, vor allem Vertreter der Haplocerataceae wie Fontanesiella und Paralingulaticeras. In den Plattenkalken der Unteren und Oberen Schiefer findet man nicht
selten Formen der Gattung Glochiceras (Untergattung Lingulaticeras; L. solenoides) mit Mündungsapophysen. Perisphinctiden mit Apophysen aber sind in den
typischen Plattenkalken extrem selten. Ein Nachweis von Marken mit Apophyseneindrücken ist mir nicht bekannt. Damit wäre diese Platte der erste entsprechende Beleg.
>> Nachlesen im Originalartikel (freier Download - open access)
>
(click on pictures to enlarge)

Abbildung 1: Die Platte mit den von links unten nach rechts oben verlaufenden Schleif- und Drehmarken eines Ammonitengehäuses. Die weißen Linien zeigen die hauptsächlichen linearen Triftrichtungen an. Abmessungen 38 x 27 Zentimeter.
Abbildung 2: Zum Vergleich eine typische Paintener Markenplatte mit Gehäuse-Rollmarken aus dem Weißjura epsilon (Oberjura; oberes Kimmeridge) von Painten ca. 20 Kilometer westlich von Regenburg, Südliche Frankenalb, Bayern. Maximal ca.37 x 27 Zentimeter. Erkennbar sind zwei um ca. 35 Grad voneinander abweichende Haupt-Strömungsrichtungen. Auf der Platte sind zahlreiche Strömungsmarken erkennbar sowie die Rollmarken von vier Ammonitengehäusen und einige undeutliche weitere. Ich erhielt die Platte im Dezember 1990 von Karin Pförringer/Regensburg. Sammlungsnummer 111290/4.

Abbildung 3: Die Markenplatte mit der eingetragenen Lage der unten nachfolgenden Detailbilder (Abb. 4 bis 10). Die schwarzen Zahlen stimmen nicht mit der Abbildungsnummer der folgenden Bilder überein, sondern korrespondieren mit der jeweils in der Bildecke unten rechts eingetragenen Zahl.
Abbildung 4: Drehmarken, dreimal um jeweils etwa 3,5 bis 4 Zentimeter versetzt. Seitlich die Eindrücke einer Apophyse - bevorzugt einseitig - leichte Schräglage des Gehäuses? Die eingetragene Kontur mit der Bezeichnung 5 entspricht Abbildung 8 und zeigt vermutlich eine Überkippung des Gehäuses in der Medianebene an ohne nachfolgende Bodenberührung des Venters (siehe Abb. 8 bis 12). Lage des Bildausschnittes siehe Abb. 3. Bildbreite 17,5 Zentimeter.

Abbildung 5: Fortführung der Mark und Richtungswechsel (auf der Platte nach oben) durch mehrfach
unterbrochenen seitlichen Schub. Am Ende dieser Achse vermutliche Überkippung des Gehäuses in der Medianachse ohne nachfolgende Bodenberührung des Venters - siehe Abb. 8 bis 12. Die Kontur mit der Bezeichnung 6 entspricht Abb. 9. Nach der Überkippung trudelte das
Gehäuse nach rechts unten; es folgen mehrfache Bodenberührungen mit Drehungen. Lage des Bildausschnittes siehe Abb.
3. Bildbreite 16 Zentimeter.
Abbildung 6: Weiterbewegung des Gehäuses, mehrfaches Auftippen der Apophysen auf den Boden und schließlich
eine ausgeprägte Aufsetzmarke knapp vor dem Plattenaußenrand. Lage des Bildausschnittes siehe Abb. 3. Bildbreite ca. 17
Zentimeter.
(click on pictures to enlarge)
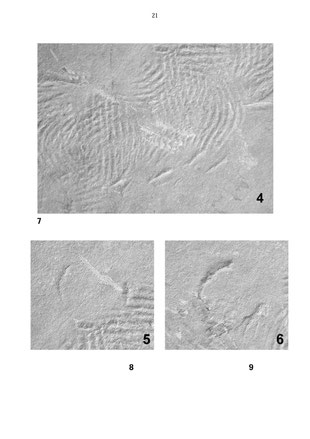
Abbildung 7: Vergrößerung der Marke im Bereich 4. Deutlich erkennbar die Drehmarken um die Vertikalachse des
Gehäuses und die Repetition der Apophyseneindrücke mit geringem seitlichem Versatz. Lage des Bildausschnittes siehe
Abb. 3. Bildbreite ca. 10 Zentimeter.
Abbildung 8 (unten links): Vergrößerung eines Mündungseindruckes nach einer Überkippung des Gehäuses, ohne
nachfolgende Boden-berührung des Venters. Siehe hierzu Abb. 10 bis 12. Lage des Bildausschnittes siehe Abb. 3 und 4.
Bildbreite ca. 4 Zentimeter.
Abbildung 9 (unten rechts): Vergrößerung eines weiteren Mündungseindruckes nach Über-kippung; auch hier
folgte keine Bodenberührung des Venters. Siehe hierzu Abb. 10 bis 12. Lage des Bildausschnittes siehe Abb. 3 und 5. Bildbreite ca. 4 Zentimeter.

Abbildungen 10 bis 12 (von links nach rechts): Entstehung der auf Abb. 8 und 9 sichtbaren Strukturen: Das
Gehäuse setzt mit dem Venter auf, schwimmt leicht auf, dreht sich in der Medianebene, setzt mit der Mündung auf, steigt auf und senkt sich etwas entfernt wieder ab.
Abbildung 13: Eine deutlich und markanter als alle anderen Marken ausgeprägte Aufsetzmarke, gefolgt von
weiteren Trudelbewegungen des Gehäuses. Lage des Bildausschnittes siehe Abb. 3. Bildbreite ca. 10 Zentimeter.
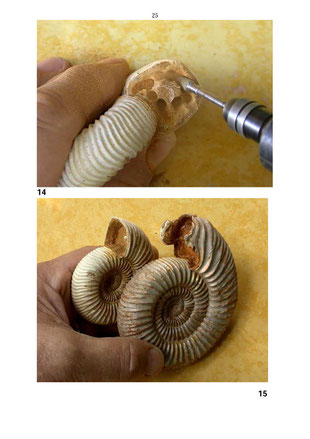
Abbildung 14: Das kleine apophysenlose Divisosphinctes-Gehäuse aus dem Oxford Madagaskars wird durch
mehrere Bohrungen mit einem Fräswerkzeug für die Aushöhlung der Gehäuseröhre vorbereitet. Dies geschah mit einem Fräswerkzeug in einer Kleinbohrmaschine.
Abbildung 15: Die beiden Divisosphinctes-Gehäuse nach Aushöhlung des Endbereichs der Gehäuseröhre.
Durchmesser des kleineren Exemplars 6,8 Zentimeter, des größeren Gehäuses 10 Zentimeter.
(click on pictures to enlarge)

Abbildung 16: Die beiden Ammoniten, locker aufgesetzt auf den plastischen Töpferton.
Abbildung 17: Die beiden Ammoniten wurden nach hinten umgelegt. Man erkennt nun die Ventereindrücke und die
Eindrücke im Mündungsbereich. Der mit geringstem Druck erzeugte Eindruck der apophysentragenden Mündung zeigt die Apophysen-Eindrücke sowie den Venterrand; bei stärkerem Druck wäre eine typisch halbkreisförmige Struktur wie auf Abb. 19, 20 und 26 entstanden,
identisch mit den Marken auf den Abb. 8 und 9.

Abbildung 18 (oben links): Geringer seitlicher Versatz mit kurzer Schleifmarke, Rückroll-bewegung auf dem
Venter, Umkehr und Bewegung der Mündung Richtung Boden, Abdruck der Mündung und wiederum Rück-bewegung (Gehäusependeln).
Abbildung 19 (oben rechts): Zurück bleibt als Mündungsabdruck eine halbkreisförmige Struktur, die identisch
ist mit den Strukturen auf den Abb. 8 und 9.
Abbildung 20: Venteraufsatz und Eindruck der Röhrenmündung beim apophysentragenden Ammonitengehäuse. Je nach
Intensität der Bodenberührung bleibt der Mündungseindruck leicht offen wie hier im Bild oder er zeigt einen geschlossenen Halbkreis wie auf Abb. 19 zu sehen.

Abbildung 21: Venter- und Mündungseindruck des apophysenlosen Gehäuses. Im Mündungs-bereich kann nur die
Venterkante eingedrückt werden. Dann wurden die Gehäuse durch Abrollen und mit achsenfixierten Drehungen auf dem Ton
bewegt. Durch Pendelbewegungen wurden die Mündungen in den Ton eingedrückt. Die Ergebnisse wurden fotografiert. Eine der Tonplatten wurde gebrannt
und als Beleg zur Markenplatte gelegt.
Abbildung 22: Das größere Gehäuse wird mit Drehbewegungen bei relativ kräftigem Druck auf der Oberfläche des
plastischen Töpfertons hin und her bewegt.
(click on pictures to enlarge)

Abbildung 23: Aufsatz-, Schleif- und Dreh-marken auf Ton.
Abbildung 24: Drehmarken und Schleifmarke (rechts) auf Ton.

Abbildung 25: Schleifmarken auf Ton.
Abbildung 26: Die als Beleg gebrannte Tonplatte mit Aufsatz-, Roll-, Schleif- und Drehmarken und
Mündungseindrücken. Plattengröße ca. 15 x 11,5 Zentimeter.
____________________________________________________________
Artikel aus dem Archiv von geomnia zu
Oenosaurus muehlheimensis
Publiziert in: Rauhut OWM, Heyng AM, Lopez-Arbarello A, Hecker A (2012) A New Rhynchocephalian from the Late Jurassic of Germany with a Dentition That Is Unique amongst Tetrapods. PLoS ONE 7(10): e46839. doi:10.1371/journal.pone.0046839
Zu Füßen der Dinosaurier: Ein jurassischer Bruder der Brückenechsen
Pressemitteilung von Oliver Rauhut & Adriana López-Arbarello, 31. Oktober 2012, published online 2012, last modified 14-01-2022
Brückenechsen werden oft als klassisches Beispiel eines „lebenden Fossils“ angesehen und gelten als letzte Überlebende einer alten Entwicklungslinie, die den moderneren Eidechsen evolutiv hoffnungslos unterlegen war. Ein neuer Fund aus dem oberen Jura (vor ca. 148 Mio Jahren) Süddeutschlands zeigt nun, dass die Brückenechsen noch zur Zeit der Ausbreitung der Eidechsen eine ungewöhnlich hohe Anpassungsfähigkeit und ökologische Vielfalt aufwiesen, was die Idee der evolutiven Unterlegenheit in Frage stellt.
Sie sieht aus wie eine “normale” Eidechse, aber gehört in Wirklichkeit zu einer alten Entwicklungslinie, die sich unabhängig von den Eidechsen entwickelt hat: Die Brückenechse, von der nur zwei Arten bekannt sind, die auf einigen wenigen kleinen Inseln vor Neuseeland leben. Mit einer Gehirnstruktur und einer Fortbewegungsweise, die oft als Intermediär zwischen Amphibien und Eidechsen bezeichnet wird, gehören Brückenechsen sicherlich zu den rätselhaftesten Reptilien und werden oft als typisches „lebendes Fossil“ angesehen, als hervorragendes Modell dafür, wie ein Urahne der Eidechsen ausgesehen haben mag. Zur Zeit der Dinosauriern, dem Mesozoikum („Erdmittelalter“) waren Vertreter der Rhynchocephalen („Schnabelköpfe“), zu denen die Brückenechsen gehören, allerdings divers und weit verbreitet. Die Frage, warum diese Gruppe im späten Erdmittelalter dem Untergang geweiht war und nur auf abgelegenenen Eilanden überlebte, schien bisher leicht zu beantworten: Mit ihrer so offensichtlich primitiven Struktur waren diese Tiere den sich in jener Zeit ausbreitenden Eidechsen und frühen Säugetieren klar unterlegen und wurden von diesen verdrängt. Waren sie dies wirklich?
Ein neuer fossiler Verwandter der Brückenechsen aus dem obersten Jura von Süddeutschland stellt diese Idee nun in Frage. In einem gerade in der wissenschaftlichen Zeitschrift PLoS One erschienenen Arbeit erhielt dieses Tier den Namen Oenosaurus muehlheimensis, zu Ehren des hervorragenden Weines von der Frankenalb und des Fundortes, dem kleinen Dorf Mühlheim, bei Mörnsheim. Oenosaurus sieht den heutigen Brückenechsen sehr ähnlich, unterscheidet sich von ihnen jedoch in der Bezahnung, die für Landwirbeltiere einzigartig ist. „Als das Stück gefunden wurde, und der Schädel nur in Gaumenansicht sichtbar war, rätselten wir alle, worum es sich wohl handeln könnte, da niemand von uns jemals eine solche Bezahnung bei einem offensichtlichen Landwirbeltier gesehen hatte“, erinnert sich Dr. Oliver Rauhut von der Bayerischen Staatssammlung für Paläontologie und Geologie in München, der Erstautor der Studie. Die Zähne von Oenosaurus bestehen aus massiven Zahnplatten, deren Struktur darauf hindeutet, dass sie zeitlebens kontinuierlich wuchsen, um damit der Abnutzung entgegenzuwirken. Ähnliche Zähne kannte man bisher nur bei Fischen, etwa bei Meerkatzen (Verwandte der Haie) und Lungenfischen. „Wir haben eine Computertomographie der Zahnplatten gemacht, und als ich die Bilder davon einer englischen Kollegin zeigte, die auf Mikrostrukturen von Fischzähnen spezialisiert ist, fand sie zunächst nichts ungewöhnliches daran – bis ich ihr sagte, dass es sich um Zähne eines Reptiles handelt“, sagt Dr. Adriana López-Arbarello, Fisch-Spezialistin der Staatssammlung und eine der Co-Autorinnen der Arbeit. „Da fiel sie aus allen Wolken und konnte es erst gar nicht glauben“, fügt sie mit einem leichten Lächeln hinzu. Diese einmalige Bezahnung bedeutet eine bisher unbekannte ökologische Anpassung bei dieser Ur-Brückenechse, die offenbar auf das Knacken hartschaliger Nahrung spezialisiert war. Rhynchocephalien haben üblicherweise eine sehr spezialisierte Bezahnung, die zum Zerschneiden von Nahrung geeignet ist und von die man bisher als limitierenden Faktor bei der Evolution der Gruppe angesehen hat. Somit zeigt Oenosaurus, dass diese Gruppe sehr viel anpassungsfähiger war, als bisher angenommen, und unterstreicht ihre große morphologische und ökologische Vielfalt im oberen Jura in Europa, kurz bevor sie hier aus dem Fossilbericht verschwinden. Dies widerspricht der gängigen Hypothese, dass Rhynchocephalen den Eidechsen und frühen Säugetieren evolutiv unterlegen waren und die Verdrängung durch diese Gruppen ausreicht, um ihren Untergang zu erklären. Stattdessen dürften Klimaveränderungen im Zusammenhang mit dem Auseinanderbrechen des Superkontinentes Pangäa eine große Rolle bei dem Untergang der Rhynchocephalen gespielt haben.
Die Überreste von Oenosaurus wurden von Roland Pöschl in Gesteinen der Mörnsheim Formation im Steinbruch am Schaudiberg bei Mühlheim gefunden. Die Steinbruchbesitzer erkannten die wissenschaftliche Bedeutung des Fundes und spendeten ihn der Bayerischen Staatssammlung für Paläontologie und Geologie in München. Die Mörnsheimer Schichten sind etwas jünger als die bekannten, darunterliegenden Solnhofener Schichten, aus denen unter anderem der berühmte Urvogel Archaeopteryx stammt. Sie sind sehr fossilreich, jedoch ist ihre Fauna viel weniger bekannt, da sie, im Gegensatz zu den Solnhofener Schichten, kaum kommerziell abgebaut werden. „Wir fangen gerade erst an, diese Gesteine zu erforschen, und das wird sicherlich noch viele weitere Überraschungen ergeben“, ist sich Alexander Heyng sicher. Der Geologe untersucht die Gesteinsabfolge am Schaudiberg und hat auch den Kontakt zwischen Steinbruchbesitzern und Paläontologen hergestellt. Ein großer Teil des Steinbruches ist der Öffentlichkeit inzwischen als Besuchersteinbruch zur Fossilsuche zugängig (http://www.besuchersteinbruch.de/). Somit kann jeder dort aktiv an der faszinierenden Entdeckung der jurassischen Lebewelt teilnehmen und die Wissenschaft in ihrer Aufgabe, die Wurzeln unserer modernen Lebewelt zu entziffern, unterstützen.

Abbildung 1: Die bis zu 50 cm lange Brückenechse, Sphenodon punctatus, die auf einigen wenigen Inseln vor Neuseeland lebt.
(Photo Dr. Helmut Tischlinger)
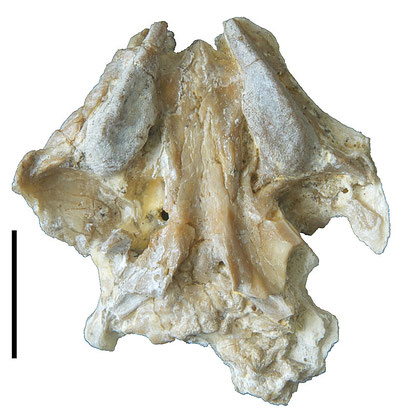
Abbildung 2: Gaumenansicht des Schädels von Oenosaurus, mit den gut sichtbaren Zahnplatten. Maßstab ist 1 cm. (Photo Krautworst)
A new taxon of sphenodontian with unusual dentition
from the Late Jurassic of Southern Germany
Oliver Rauhut, Alexander M. Heyng & Andreas Hecker, published online 25-02-2010, last modified 14-01-2022
Although the only recent representative of the Sphenodontia, the genus Sphenodon, is often used as a “classic” example of a living fossil, research in the recent decades has shown that this clade was widespread and both taxonomically and ecologically diverse during the Mesozoic. All sphenodontians but the most basal forms are characterized by a special, acrodont dentition, in which a juvenile dentition is retained throughout the ontogeny, and new teeth are added posteriorly in the jaws. Recently, the partial skull and mandibles of a new taxon of sphenodontian have been discovered in the marine Tithonian Mörnsheim Formation in Bavaria, southern Germany. Although the skull roof and braincase are fragmentary, the palate and lower jaws are excellently preserved. The most conspicuous character, however, is the very unusual dentition, which consists of large lateral tooth plates in the maxilla and dentaries. This character strongly indicates a durophagous diet in the new taxon. The tooth plates are formed by the fusion of numerous, small, pencil-like teeth, very unlike the typical tooth shape in sphenodontians, and there is no distinction between a juvenile tooth row and additional teeth. Nevertheless, phylogenetic analysis indicates that the new taxon is well nested within sphenodontians that have the typical type of dentition of this clade; thus, its tooth plates should be derived from an acrodont dentition with retention of a juvenile dentition and posterior aggregation of additional teeth. This indicates a surprisingly high evolutionary plasticity in the dentition of derived sphenodontians. A possible evolutionary origin for this dentition might be the heterochronic retention and subsequent modification of small, more conical teeth, as they are found in hatchlings of the recent Sphenodon.

Figure 3: Detailed drawing in ventral view of the fragmentary preserved skull of the new sphenodontian taxon.
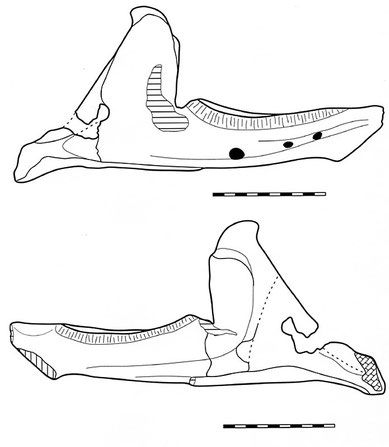
Figure 4: Sketch of the excellently preserved lower jaw.
Beschreibung einer neuen Echsenart
Published online 21-01-2009 by Andreas Hecker, last modified 14-01-2022
Im April 2008 wurde im Steinbruch Krautworst am Schaudiberg ein kleiner Schädel und die dazugehörigen Unterkiefer gefunden. Es handelt sich dabei um eine bisher unbekannte Art. Das Fossil kann den Rhynchocephalen zugeordnet werden, die schon seit langem mit vielen Gattungen aus den Solnhofener und Mörnsheimer Schichten bekannt sind (siehe z.B. FRICKINGER1994). Besonders zu nennen sind hier die Gruppe der Pleurosaurier und Gattungen wie Homoiosaurus und Kallimodon. Als einziger heute noch lebender Vertreter gilt Sphenodon punctatus GRAY aufgrund seiner primitiven Merkmale als lebendes Fossil (ROMER 1956).
Lange Zeit wurde angenommen, die Rhynchocephalia seien eine hochkonservative Gruppe, lediglich die Pleurosaurier wären mit ihrer sehr weitgehenden Anpassung an die aquatische Lebensweise (COCUDE-MICHEL 1963) stärker spezialisiert. Neuere Funde aus dem Oberen Jura Nord-Afrikas und der Kreide Süd-Amerikas belegen jedoch starke Spezialisierungen an verschiedene Lebensweisen (THROCKMORTON, HOPSON, PARKS 1981; REYNOSO 1997; REYNOSO 2000; APESTEGUJA 2005; JONES 2008).
Die stark abgewandelte Bezahnung des neuen Fundes aus Mühlheim unterstreicht die ungeahnte Anpassungsfähigkeit der Rhynchocephalier. Der Fund wird wissenschaftlich bearbeitet (Publikation in Vorbereitung).
Zitierte Literatur
- REYNOSO, V. (1997): A ”BEADED'' SPHENODONTIAN (DIAPSIDA: LEPIDOSAURIA) FROM THE EARLY CRETACEOUS OF CENTRAL MEXICO, - Journal of Vertebrate Paleontology V. l7(1), pp. 52-59
- REYNOSO, V. (2000): AN UNUSUAL AQUATIC SPHENODONTIAN (REPTILIA: DIAPSIDA) FROM THE TLAYUA FORMATION (ALBIAN), CENTRAL MEXICO, - Journal of Paleontology V. 74(1), pp. 133–148
- FRICKINGER, K. A. (1994): DIE FOSSILLIEN VON SOLNHOFEN, - Goldschneck Verlag
- COCUDE-MICHEL, M. (1963): LES RHYNCHOCEPHALES ET LES SAURIENS DES CALCARIES LITHGRAPHIQUES (JURASSIQUE SUPERIEUR) D´ EUROPE OCCIDENTALE, - Nouvelles Archives du Museum d´histoire naturelle du Lyon, V. 7, 1963, pp. 1-187
- THROCKMORTON, G. S.; HOPSON, J. A.; PARKS, P. (1981): A REDESCRIPTION OF TOXOLOPHOSAURUS CLOUDI OLSON; A LOWER CRETACEOUS HERBIVORUS SPHENODONTID REPTILE, - Journal of Paleontology, V. (3), pp. 586 - 597
- APESTRGUJA, S. (2005): A LATE CAMPANIAN SPHENODONTID (REPTILIA; DIAPSIDA) FROM NORTHERN PATAGONIA, - C. R. Palevol, V. 4, pp. 663 – 669
- JONES, M. E. H. (2008): SKULL SHAPE AND FEEDING STRATEGY IN SPHENODON AND OTHER RHYNCHOCEPHALIA (DIAPSIDA: LEPIDOSAURIA), - Journal of Morphology, V. 269, pp. 945 - 966
- ROMER, A. S. (1956): OSTEOLOGY OF THE REPTILES, - University of Chicago Press, 1956
____________________________________________________________
Aus dem Archiv der Zeitschrift Documenta naturae (Band 192-4, 2013)
A rare new demosponge from the Solnhofen Lithographic Limestone
(Upper Jurassic, Bavaria, Germany)
J. Reitner*
*Prof. Dr. Joachim Reitner, Geobiology, GZG, University of Göttingen, Goldschmidtstr. 3, 37077 Göttingen, Germany
Abstract: From the Lower Tithonian of the Eichstätt realm a new exceptional demosponge taxon is described. The new taxon Hruodospongia lithographica n. gen. n. sp. is related to the Halichrondrida/Axinellida based on the spicule arrangements and architecture. The sponge grew on a stem of unknown origin and was probably drifting for a while from a shallower, oxygen-rich environment before sinking down in the oxygen-depleted deeper water of the Eichstätter "Wanne" (lagoonal basin). Sponges are rare within the typical Plattenkalk facies, because the oxygen-poor benthic environments which did not allow the growth of sessile benthos which were transported from carbonate platforms nearby. Discussed is also the paleoecology and systematic relationship of further dubious demosponges found in the Plattenkalk facies.
Key words: Upper Jurassic, Tithonian, Porifera, demosponges, Solnhofen Lithographic Limestone, Bavaria, Germany.
Introduction: Sponges are usually very common in the upper Jurassic and often form a special reef-type facies (e.g. Leinfelder 1993). Normally most sponges prefer clear, oxygen rich oligotrophic water which is characteristic for most reef facies. The Kimmeridgian and Tithonian sponge build-ups were dominated by lithistid demosponges cemented by microbial crusts. Within the older Oxfordian sponge reefs, hexactinellid sponges have often played the dominate role. The lower Tithonian Plattenkalk facies is a typical restricted lagoonal facies embedded between sponge and coral reefs of the vast reef belt of the northern margin of the alpine ocean system. The typical Tithonian Plattenkalk facies (Malm zeta 2b) exhibits only few occurrences of benthic organisms (ophiurids, polychaetes, bivales, etc.) and they are restricted to certain lagoons ("Wannen"), e.g. from Pfalzpaint, Zandt, and some others (Röper et al. 1999, 2001; Keupp & Schweigert 2012) which probably had short-time oxygen enriched water masses. The lagoonal basins of the southern Franconian Alb exhibit two major sedimentological-stratigraphic events, the so-called "Krumme Lagen", the lower "Trennende Krumme Lage" which separates the lowermost Tithonian (zeta 2a) from the central Plattenkalk series (zeta 2b) and the "Hangende Krumme Lage" from the "Mörnsheimer Schichten" (zeta 3) (Barthel 1978; Viohl 1998; Kölbl-Ebert et al. 2005). These up to meters thick layers were probably caused by earth quakes strongly synsedimentarily folded and sometimes bearing shallow water organisms like sponges from the surrounding reefs. These events could mixed up the stagnant lagoonal waters with oxygen. Sponges are very rare and only two taxa have up to now been described, the hexactinellid Ammonella quadrata Walter 1904 (Keupp & Mehl 1994) and the demosponge Neochoiaella frattigianii Keupp & Schweigert 2012. The autochthonous nature of these sponges is questionable, it is more convincing that they were transported from a shallower oxygenated environment.
>> Weiterlesen im Originalartikel (freier Download - open access)
>

Fig.1. Holotype of Hruodospongia lithographica n.gen. n.sp. Collection of the Geoscience Museum Göttingen GZG.INV. 78089. Arrow shows stick of unknown origin on which the sponge was grown.
Fig.2. Bundles of diactine spicules (arrow) of the dermal layer. Former siliceous spicules are preserved either in oxidized pyrite crystals (brown dark spots) or in calcite.
(please click on figures to enlarge)
Fig.3. Thick dermal spicule layer with abundant spicules preserved in knobby calcite (red arrow). Within the former sponge tissue abundant star-shaped calcite crystal aggregates are shown, probably an early diagenetic phase of tissue preservation (black arrow).
Fig.4. Same as Fig.3 with clear plumose-like arranged dermal spicules preserved in calcite.
Fig.5. Reticulately arranged choanosomal diactines (arrows) preserved in iron hydroxides, probably oxidized pyrite.
(please click on figures to enlarge)

Fig. 6. Codites ("Phyllothallus") elongatus
Sternberg 1833. These types of fossils are relatively common and traditionally interpreted as brown algae. Dietl & Schweigert (1999, 2001) re-interpreted these fossils as "Weichschämme- soft
sponges" because they detected large kidney-shaped microscleres (rhax, sterraster, selenaster). "Weichschwämme" is definitely not a good description of this sponge type. The strong dermal layer
caused by a dense crust of microscleres gave the sponges a hard and stable character, as known from modern relatives. The term "soft sponges" is often used for non-spiculated keratose-type
sponges and should be restricted to them. The taxonomic position is unclear, since different taxa of demosponges bear these types of microscleres and megascleres are also not known. Anatomically
best coincidences are seen with the taxon Chondrilla grandistellata Thiele 1900, a hadromerid sponge with a prominent dermal layer of large euasters. However, the kidney-shaped
microscleres of Codites ("Phyllothallus") elongatus resemble more the selenasters of the hadromerid Placospongia.
Fig.7. Detailed view of the dermal layer of Codites ("Phyllothallus") elongatus exhibiting abundant large kidney-shaped microscleres (red arrow) and abundant oscula pores (black arrow), which finally prove the sponge nature of these fossils.
(please click on figures to enlarge)
Published online 10-01-2022 by A. M. Heyng
____________________________________________________________
Historische Abgüsse von Plattenkalk-Fossilien
Published online 02-07-2021 by A. M. Heyng, last update 12-01-2022


















Beim Begutachten der ehemaligen Lehrsammlung des Königlichen Schullehrer-Seminars in Eichstätt fanden sich im "Sammelsurium" zwischen den Originalen auch Abgüsse von Plattenkalk-Fosssilien - in arg lädiertem Zustand, aber immerhin, derartige historische Belege sind nicht so häufig zu finden und definitiv aufhebenswert!
Wann die Abgüsse letztendlich gefertigt wurden, ist nicht bekannt (ab 1835 ist möglich, wahrscheinlich aber viel später). Deutlich erkennbar ist jedoch, dass sich - neben der Verwendung anderer
Materialien - auch die Abgusstechnik und die Art der Koloration bis heute stark weiterentwickelt hat - heutzutage sind Original und Abguss (wenn man es will) mit dem bloßen Augen kaum mehr
unterscheidbar.
Die hier im Originalzustand bei der Bergung abgebildeten historischen Abgüsse (alle ohne Inventar-Nummer) werden aufbewahrt in der wissenschaftlichen Sammlung amh-Geo.
____________________________________________________________
"Alte Stücke" aus historischen Plattenkalk-Sammlungen
Published online 30-10-2020 by A. M. Heyng

(click to enlarge)
"Athrotaxites princeps P XII 2" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk138-18, wiss. Slg. amh-Geo)


(click to enlarge)
"Athrotaxites Frischmanni P. XII 3" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk139-18, wiss. Slg. amh-Geo)

(click to enlarge)
"Comatula (Saccocoma) pectinata X 5" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk467-18, wiss. Slg. amh-Geo)


(click to enlarge)
"Palinurina intermedia V8.2" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk203-18, wiss. Slg. amh-Geo)


(click to enlarge)
"Glyphea V2" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt, im
Originalzustand (unpräpariert).
(Inv.-Nr. pk263-18, wiss. Slg. amh-Geo)


(click to enlarge)
"Koelga V16" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk393-18, wiss. Slg. amh-Geo)


(click to enlarge)
"Amonites IX" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk401-18, wiss. Slg. amh-Geo)

(click to enlarge)
"Aptychus latus IX" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk432-18, wiss. Slg. amh-Geo)


(click to enlarge)
"Leptolepis? IV 20" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk426-18, wiss. Slg. amh-Geo)


(click to enlarge)
"Gyrodus IV 26" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk186-18, wiss. Slg. amh-Geo)

(click to enlarge)
"Leptolepis macrolepidotus IV 20" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk524-20, wiss. Slg. amh-Geo)

(click to enlarge)
"Lumbricaria VIII 2" aus der ehemaligen Lehrsammlung des "Königlichen Schullehrer-Seminars" in Eichstätt.
(Inv.-Nr. pk392-18, wiss. Slg. amh-Geo)
____________________________________________________________
New Paper: Muensterelloidea and the evolution of octopods

(please click to enlarge)

Holotype of Muensterella spinosa sp. nov. (BSPG 1964XXII 103a) from the lower Tithonian Moernsheim Formation (hybonotum Zone, moernsheimensis Subzone) of Daiting.
Fuchs, D., Iba, Y., Heyng, A., Iijima, M., Klug, Ch., Larson N. L., & Schweigert, G. 2019. THE MUENSTERELLOIDEA: PHYLOGENY AND CHARACTER EVOLUTION OF MESOZOIC STEM OCTOPODS. Papers in Palaeontology, 1–62. https://doi.org/10.1002/spp2.1254
Abstract: The Muensterelloidea is a superfamily of teudopseid octobrachians with a posteriorly patella‐shaped gladius. A morphometric comparison based on 148 muensterelloid gladii has yielded five new species accommodated in three new genera: Engeseriteuthis arcuatus gen. et sp. nov., Muensterella jillae sp. nov., Muensterella spinosa sp. nov., Tyrionella fauseri gen. et sp. nov. and Muensterellina johnjagti gen. et sp. nov. Cretaceous taxa ‘Tusoteuthis’ cobbani and ‘Muensterella’ tonii are re‐combined and placed within the genus Enchoteuthis. We introduce categories for gladius proportions applicable for both muensterelloid and non‐muensterelloid octobrachian gladii. A 2D‐landmark analysis including 64 muensterelloid and non‐muensterelloid gladii statistically confirms that the Muensterelloidea possess the smallest median field sizes of all Mesozoic gladii. We consider the lateral field‐dominated ‘gladius’ of the family Patelloctopodidae (Patelloctopus, Pearceiteuthis) to be vestigial (i.e. shorter than the mantle length) suggesting that the Patelloctopodidae are the last shared ancestors of incirrate and cirrate octopods. According to a phylogenetic analysis based on 31 gladius characters, the Muensterelloidea mainly consists of the ‘Muensterella‐Enchoteuthidae’ and the ‘patelloctopodid’ clade. Ancestral character state reconstructions suggest that an increasing posterior growth front is accompanied by a continuous decrease of the median field length. This milestone in the evolution of the octopod gladius vestige occurred between the Early and Middle Jurassic. The benthic life style of incirrate octopods (including Cretaceous palaeoctopodids) was adopted by Jurassic Patelloctopodidae, which arose from nectonic to nectobenthic teudopseid ancestors. There is currently no evidence to suggest a pelagic origin for benthic octopods.
Key words: Coleoidea, Octopoda, gladius vestige, phylogeny, shell evolution.
Published online 16-01-2020 by A. M. Heyng
____________________________________________________________
(please click to enlarge)
First record of discinid brachiopods from Late Jurassic Plattenkalk deposits of southern Germany
Autoren: Mike Reich, Alexander M. Heyng, Bork Ilsemann, Alexander Nützel & Tanja R. Stegemann
Präsentation: 88. Jahrestagung der Paläontologischen Gesellschaft, 26.-30. März 2017, Münster
Introduction
The Kimmeridgian and Tithonian Plattenkalk deposits of southern Germany (e.g., Solnhofen, Eichstätt, Pfalzpaint, Painten, Brunn, Wattendorf) are famous for their diverse and exceptionally preserved fauna and flora. So far, there are around 1,800 species, mostly macrofossils, reported from these strata. However, with the help of non-applied or hitherto unused micropalaeontological methods, new (or so far unrecorded) species or groups can be detected.
In the present study we report the first find of organophosphatic-shelled brachiopods (Linguliformea: Discinoidea) from the Late Jurassic Plattenkalk deposits of southern Germany.
Fossil and modern Discinida
Linguliform brachiopods were major components of marine assemblages from the Early Cambrian onwards, but their evolution and palaeogeography during the Jurassic–Cretaceous is poorly understood, as a result of a sparse fossil record.
Discinid brachiopods are epifaunal, fixo-sessile invertebrates, common in some Recent environments (intertidal) and attached by a highly muscular pedicle to various hard substrates. The discinid pedicle acts as an anchor, and it supports the weight of the shell and holds it in relative position to the substrate. The ventral valve is always oriented toward the substratum (Fig. A), a feature that discinids share with craniids and articulate thecideid brachiopods. Modern discinids (solitary or known in clusters; Fig. B) have a cryptic mode of life on the underside of pebbles or attached to larger boulders embedded in soft substrate.
In the fossil record (since the Ordovician), discinids are also known attached to articulate brachiopods, bivalves, cephalopods, and conulariids.
Importance of the material
As already stated this is the first record of organophosphaticshelled brachiopods from Late Jurassic Plattenkalk deposits of southern Germany. Summarized, our data reveal a linguliform brachiopod diversity hitherto unknown from Jurassic Plattenkalk Lagerstätten. The new recorded data will help to develop a more detailed palaeobiodiversity as well as palaeoecological interpretation of Late Jurassic Plattenkalk deposits in southern Germany. The discovery of a diverse discinid brachiopod fauna offers in part new insight into palaeoenvironmental patterns and former benthic habitats of the Late Jurassic Solnhofen Archipelago. The applied micropalaeontological methods in isolating microscopic/mesoscopic invertebrates is seen to be promising for the study of neglected or overlooked fossils in general.
Results
Discinid brachiopod shells (especially ventral valves) are generally rare in the Mesozoic/Cenozoic fossil record, therefore they have often been taken to indicate a low preservational potential. The here presented organophosphatic brachiopod material is based on >100 (microscopic / mesoscopic) isolated valves (rock disintegration by using buffered acetic acid) from the Early Tithonian Mörnsheim Formation (Hybonotum Zone: Moernsheimensis Subzone, ~149 Ma; Fig. C) of two sections in Mühlheim near Solnhofen, Upper Bavaria, Germany.
Three species of the Discinidae (Figs. D-G) can be identified from our strata, including a new genus and species (related to Discinisca and Discradisca). In
addition, we documented the associated macro-, meso-, and microfauna (Reich et al. in prep.) of the studied sections. All recorded discinid brachiopods are associated with sponges
(Hexactinellida, Demospongea), bivalves and other invertebrates and microfossils.
Acknowledgements: We are indebted to Marlena Olbrys (formerly LMU Munich) for technical help. Furthermore, we
would like to thank the owners and operators of the Mühlheim quarries, Ulrich Leonhardt, Roland Pöschl, and Uwe Krautworst, for logistics and support.
Published online 19-12-2017 by M. Reich, A. M. Heyng, B. Ilsemann, A. Nützel & T. R. Stegemann
________________________________________________________________________________________________________________________________________________________________________
__________________________________________